1、能是做功的本领
能的定义就是做功的本领,做功就是指让物体移动位置。生物体内做的功种类很多,都是需要能量的。没有能,生物就不可能存活。
能量可以分为动能( kinetic energy)和势能( potential energy)两种形式。
动能 | 势能 | |
概念 | 运动的能量 | 物体由于所在位置或本身的排列而具有的能量 |
举例 | 热、电和光都是动能 | 活细胞中做各种功的化学能,是活的生物体内最重要的能量形式 |
2、热力学定律
能量可以从一种形式转变为另一种形式。
热力学( thermodynamics)是研究所有物体中能量转化规律的科学。在讨论能量转化时,我们把所研究的物体或物体的集合体称为体系( system ) ,而把宇宙的其余部分统称为环境。
开放体系是所有生物的特点。
- 热力学第一定律认为宇宙中的总能量是固定不变的。这条定律也称为能量守恒定律。能量既不能被创造,也不能被消灭,只能从一种形式转变为另一种形式。
- 热力学第二定律说明能量的转变导致宇宙的有序性减低而无序性增加。一个特定体系的有序性要增加,其环境的无序性便必须增加。细胞中能量转化的效率不可能是100%。在所有能量转换过程中,总有一部分能量以热的形式逃逸,这是不可避免的。
3、吸能反应和放能反应
化学反应可分为吸能反应(endergonic reaction)和放能反应( exergonic reaction)两大类,细胞中发生的化学反应也不例外。
吸能反应 | 放能反应 | |
概念 | 指反应产物分子中的势能比反应物分子中的势能多 | 其产物分子中的化学能少于反应物分子中的化学能 |
举例 | 光合作用是生物界最重要的吸能反应。 | 木材的燃烧;细胞呼吸 |
每一个活细胞中都要发生千百种放能反应和吸能反应,所有这些反应总称为细胞代谢( cellular metabolism)。 | ||
放能反应与吸能反应之间,有一种能量传递的关系,这种关系是通过纽带——ATP而发生的。 | ||
4、ATP是细胞中的能量通货
ATP( adenosine triphosphate,腺苷三磷酸)是一种核苷酸,其中有3个磷酸基团。ATP分子由一个戊糖(核糖)、一个含氮碱基(腺嘌呤)和3个磷酸根组成,其中一个磷酸根连接在糖分子上,其余两个则相继连接。这两个磷酸键比较不稳定,是焦磷酸键,称为高能磷酸键,以“~”表示。这是因为这种键水解时会释放较多的能量,比普通磷酸键的多。

ATP发生水解时,形成ADP并释放一个磷酸根,同时释放能量。这些能量在细胞中就会被利用,肌肉收缩产生的运动,神经细胞的活动,生物体内的其他一切活动利用的都是ATP水解时产生的能量。
ATP在细胞中易于再生,所以是源源不断的能源。这种通过ATP的水解和合成而使放能反应所释放的能量用于吸能反应的过程称为ATP循环。因为ATP是细胞中普遍应用的能量的载体,所以常称之为细胞中的能量通货( energy currency )。

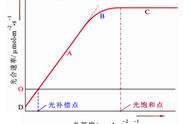
1、酶降低反应的活化能
酶( enzyme)是一种生物催化剂( biocatalyst), 绝大多数酶都是蛋白质。它能加速生物体内化学反应的进行,但在反应前后并不发生变化。
任何一个分子要发生化学反应,都必须先被活化,即增加能量。正是由于分子发生化学反应需要活化能,生物体内许多化学性质活泼的物质才能保持稳定。比如ATP是很易被水解的,但由于水解需要ATP被活化,所以在水很丰富的环境(细胞)中,ATP仍能保持稳定。ATP的水解只有在专一的酶——ATP酶( ATPase)的催化下才易于发生。
酶能降低反应的活化能( activationenergy)。
酶分子是蛋白质,每种蛋白质都有特定的三维形状,而这种形状就决定了酶的选择性。
- 酶所催化的反应中的反应物称为底物( substrate),酶只能识别一种或一类专一的底物并催化专一的化学反应。
- 所以细胞中发生的所有反应需要许多种不同的酶催化。酶只催化特定底物的特定反应,这种性质称为酶的底物专一性( substrate specificity )。
- 酶之所以有这种专一性,是因为酶会与其底物结合,只有当底物与酶分子结合以后,才会变成产物。酶分子中只有一个小的局部与底物分子结合,这一小的局部就称为酶的活性部位( active site)。
- 酶所以有专一性是因为其活性部位只适于结合一种或一类底物分子。
酶的作用仍需要整个酶分子,因为如果整个分子有损伤,活性部位也会发生变化,不起作用了。
2、多种因素影响酶的活性
因素 | |||
温度 | 只有在最适温度下酶活性才最高。 | ||
pH | 一般酶的最适pH为6~8 ,接近于中性。在此范围之外,酶的活性就较低,整个细胞的正常功能也被破坏。 | ||
盐的浓度 | 盐浓度太高会干扰酶分子中的某些化学键,从而破坏其蛋白质结构,也就是使其活性降低。只有极少数种类的酶能耐受极高的盐浓度。 | ||
辅因子 | 许多种酶的正常活动还需要非蛋白质成分的参与。 辅因子可能是无机物; 也可能是有机物,有机的辅因子则称为辅酶( coenzyme)。 | ||
酶的抑制剂 | 竞争性抑制剂 | 竞争性抑制剂是与酶的正常底物相似的化学物质,它与底物分子竞争酶的活性部位。 | |
非竞争性抑制剂 | 非竞争性抑制剂并不占据活性部位,它与酶分子结合的部位不是活性部位。但它的结合却使酶分子的形状发生了变化,而使得活性部位不适于接纳底物分子。 | ||
酶的抑制剂有重要的应用价值,某些*虫剂和抗生素就是酶的抑制剂,例如,马拉硫磷是乙酰胆碱酯酶的抑制剂,这种酶在神经系统的活动中非常重要。在马拉硫磷的作用下,神经细胞对信号的传递被抑制,造成昆虫死亡。 青霉素抑制细菌细胞中合成细胞壁的酶,所以能够阻止病菌的增殖,人体细胞没有细胞壁,所以青霉素对人无害。 | |||
3、核酶
科学家发现细胞中与RNA 分子有关的某些反应却是由RNA本身所催化的,而不是由蛋白质催化的。因此,不仅蛋白质是生物催化剂, RNA 也是生物催化剂。于是给这种RNA催化剂起了一个名字“核酶”( ribozyme)。
现在可能至少有两类核酶。
- 一类是催化分子内的反应,即分子的一部分与另一部分反应,例如 RNA的一段在该分子内改换位置。这时RNA分子本身既是底物﹐又是催化剂。这种RNA分子具有特定的折叠形式。
- 另一类核酶则催化分子间的反应,就是催化别的分子发生反应而作为核酶的RNA分子在反应前后无变化。
生物膜是一种超分子( supramolecular)结构,就是由多分子形成的一种有序的组织。
本节将讨论生物膜通过调节穿过细胞边界的转运物质的能力。
1、膜的选择透性源于其分子组成
决定选择透性的因素有二:脂双层和转运蛋白。
脂双层:亲脂性。烃类、二氧化碳和氧能溶于脂双层中,所以易于透过质膜。
转运蛋白(膜内在蛋白):使得亲水的离子和极性分子也能被运过脂双层的中心。许多重要的亲水分子,如葡萄糖、氨基酸等能透过脂双层,避免了与膜中亲脂部分的接触。
- 有时,转运蛋白的专一性非常强,例如,人血液中的葡萄糖经过专一的转运蛋白进入肝细胞,但是果糖(葡萄糖的异构体)却不能由此进入。
两种转运方式:被动转运( passive transport)和主动转运( active transport)。
被动转运
2、被动转运是穿过膜的扩散
扩散( diffusion)是分子因其所带动能自由运动而造成的。每一个分子的运动是随机的,然而一群分子的运动却是有方向的。
扩散的方向决定于扩散物质的浓度梯度( concentration gradient)。
- 分子或离子总是从高浓度区域向低浓度区域移动,降低其自由能。离子不仅顺其浓度梯度而扩散,而且也顺其电化学势梯度而扩散。
- 这种顺浓度梯度而穿过膜扩散的作用就是物质的被动转运。
- 被动转运是物质出入细胞中常见的现象。
当一种溶液中有多种溶质存在时,某种物质穿过膜的扩散仅与该物质的浓度梯度有关,而与其他任何溶质的浓度均无关。
被动转运虽是常见的现象,但生物膜(包括质膜)是有选择透性的,所以并不是任何顺浓度梯度存在的物质都能顺利地穿过膜。
3、渗透是水的被动转运
两种溶质相同而浓度不同的溶液,相比较而言,
- 浓度较高的称为高渗溶液;
- 浓度较低的则称为低渗溶液;
- 溶质浓度相等的溶液则称为等渗溶液。
(1)无壁细胞的水分平衡
当动物细胞被放在等渗的环境中时,既不吸收水分,也不丢失水分,细胞处于正常状态。
- 许多海生无脊椎动物与海水是等渗的。大多数陆生动物的细胞周围的液体与细胞中的液体也是等渗的。
若将细胞放在高渗溶液中,则细胞丢失水分而皱缩,此时细胞大概会死亡。
在低渗溶液中,则水分进入细胞的速率大于由细胞中出去的速率,细胞也会膨胀而破裂。
- 许多生活在低渗或高渗环境中的动物都有特殊的渗透调节的适应以控制水分平衡。例如,草履虫生活在低渗的池水中。草履虫的质膜对水的透性较小,这解决了水分不断进入细胞的问题。此外,草履虫还有一个伸缩泡,其作用是将水排出细胞的排水泵。
(2)有壁细胞的水分平衡
植物﹑藻类、细菌和真菌的细胞都有细胞壁。
- 这些细胞在低渗溶液中是膨胀的( turgid ) ,
- 在等渗溶液中是萎蔫的( flaccid ),
- 而在高渗溶液中则发生质壁分离( plasmolysis ),即细胞质皱缩而与细胞壁分开。
渗透现象是集流和扩散的综合结果。
- 集流是指浓度差或压力差所造成的一群离子或分子的集体流动。
- 渗透作用决定于这两种推动力,把它们概括在一起的概念就是水的化学势梯度。
水势是一个体系中水的化学势。
- 对于植物细胞而言,细胞的水势是由渗透势和压力势组成的。
- 渗透势决定于细胞液的总浓度,总是负值。
- 压力势来源于细胞壁的压力,总是正值。
4、专一的蛋白质使被动转运更容易
亲水性物质是通过膜上有许多种转运蛋白帮助它们通过脂双层。这种现象称为易化扩散( facilitated diffusion) 。
转运蛋白分两类:载体蛋白( carrier protein)和通道蛋白( channel protein)。
- 转运蛋白是分子或离子穿过膜的“走廊”,存在于膜中的各种通道蛋白为某种分子或离子提供一个亲水的通道,使得它们极易通过膜。水通道蛋白( aquaporin)就是动物、植物细胞膜上通道蛋白的一个代表。
- 载体蛋白,例如葡萄糖的载体蛋白。载体蛋白在与被转运的离子结合后会发生形状上的变化,将离子从膜的一侧运到另一侧。
- 通道蛋白的作用只是加速离子的运动,方向只能是顺浓度梯度。
人的胱氨酸尿症,就是因为缺少了将胱氨酸和其他氨基酸运过肾脏细胞的质膜的转运系统而引起的。正常的肾脏细胞会从尿中重新吸收这些氨基酸而使之进入血流,但此病的患者的肾脏中却积累这些氨基酸并形成晶体,于是产生肾结石。
主动转运
5、主动转运是逆浓度梯度的转运
易化扩散只是提供了一个便利的扩散通道,不能改变转运的方向。
有些转运蛋白能使溶质逆浓度梯度而被转运,从低浓度一侧穿过质膜而到达高浓度一侧,细胞必须提供其代谢能。这种跨膜转运称为主动转运( activetransport)。
主动转运使得细胞维持其内部某种小分子或离子的浓度高于其环境中的浓度。
主动转运是由埋在质膜内的专一的蛋白质完成的。给主动转运供应能量的是ATP。
- ATP推动主动转运的一种方式就是其末端的磷酸根直接转移给转运蛋白。磷酸基团的转移使得转运蛋白改变其构象(形状),使结合在其上的溶质发生穿过膜的转运。
- 例如:钠–钾泵( sodium - potassium pump)。它将钾离子泵入钾离子浓度已经很高的细胞质内,而将钠离子泵至细胞外。这种泵(转运蛋白)在两种构象中反复变换,每将3个Na 运出细胞,便将2个K 运入细胞。ATP将转运蛋白磷酸化,从而使之改变构象。

有些离子泵会产生跨膜的电势,就是使膜的两侧带有正、负两种不同的电荷。由于质膜内外两侧阴、阳离子的分布不均,所以质膜的胞内侧带负电荷,而胞外侧带正电荷。所以膜电势有利于阳离子穿膜进入细胞而阴离子被运出细胞。
所以推动离子跨膜扩散的因素有二:浓度梯度和膜电势。这两个因素合在一起称为电化学势梯度( electrochemical gradient)。
使膜两侧产生电压的转运蛋白称为生电泵( electrogenic pump)。
- 动物细胞中的生电泵:钠–钾泵。将3个Na 泵出时将2个K 泵入。每泵动一次,就有一个正电荷从细胞质中被转移至胞外,这一过程以电压的形式贮存了能量。
- 植物细胞和真菌的生电泵:质子泵。它将质子(氢离子)主动运出细胞。质子泵利用ATP作为能源,以H 的形式将正电荷运至细胞以外,造成膜两侧的电荷分离。由此形成的膜两侧的电位差和H 浓度梯度代表一种能源,可用于推动协同转运( cotransport)。
协同转运是指专一转运一种溶质(以ATP为能源)的泵又间接地推动其他电解质的主动转运。
- 例如植物细胞利用质子泵所产生的H 浓度梯度来推动糖类、氨基酸等养分被吸收进入细胞的活动。ATP推动的质子泵将H 泵出细胞,当H “漏入”细胞时,蔗糖可以逆浓度进入细胞。这种使H -蔗糖协同转运的蛋白质只有在蔗糖分子与H 共同被转运时,蔗糖才能逆浓度梯度而被转运。植物利用这种H -蔗糖协同转运的机制将光合作用产生的蔗糖运至叶脉的专门的细胞中,然后通过维管组织将蔗糖分配到非光合组织(如根)中去。