原核生物(上)

原核生物(英文:prokaryote)是通常由单一原核细胞形成的生物。相对于真核细胞,原核细胞一般没有细胞内膜、没有核膜包裹的成型细胞核,细胞内无染色体,DNA链未螺旋化,并以游离的形式存在于细胞质中,细胞质内也无任何有膜的细胞器(如线粒体或叶绿体)。有些分类学者将原核生物归于原核生物域(Prokaryota),但现行的三域系统不采此说,而是将古菌域和细菌域的生物视为原核生物,原核生物本身不作为生物分类的层级。
大部分原核生物为单细胞生物。根据《伯杰氏细菌鉴定手册》,原核生物分为四大类,“有细胞壁的革兰氏阴性真细菌”,“有细胞壁的革兰氏阳性真细菌”,“无细胞壁的真细菌”,“古细菌”。环境中常见的原核生物有细菌、放线菌、古细菌、螺旋体、衣原体、支原体、立克次氏体和蓝细菌等光合性细菌。
Prokaryota亦拼写为"procaryotes-ß",一语来自于karyon,意思是核。pro指演化出完整细胞核之前,没有完整之细胞核。原核细胞细胞质内遗传区称核区(nucleoid)。原核细胞进行复制DNA及分割的过程,称为细胞分裂。
1.特征(1)鞭毛鞭毛是很多单细胞生物和一些多细胞生物细胞表面像鞭子一样的细胞器,用于运动及其它一些功能。在三个域中,鞭毛的结构各不相同。细菌的鞭毛是螺旋状的纤维,像螺丝一样旋转,属于生命系统中的旋转运动。古菌的鞭毛表面上和细菌的类似,但很多细节不同,和细菌的鞭毛可能也不是同源的。真核生物,比如动物、植物、原生生物细胞的鞭毛是细胞表面结构复杂的突出物,像鞭子一样来回抽打。
①类型三种类型的鞭毛已经被区别:细菌的,古菌的,和真核生物的。
这三种类型之间的主要区别概括如下:
细菌鞭毛是螺旋细丝,每一个在其基部带有旋转马达可以顺时针或逆时针转动。它们提供了几种细菌运动性中的两个运动。
古菌鞭毛(Archaella)是表面上类似于细菌鞭毛,但是在许多细节上是不同的,并认为是非同源。
真核生物的鞭毛:植物,动物和原生生物的细胞的复杂的细胞凸出物像鞭子一样来回的抽打。
②细菌鞭毛不同种类的细菌有不同的数目的鞭毛。Monotrichous细菌有一个单一的鞭毛(如霍乱弧菌)。 Lophotrichous细菌有多种鞭毛设在细菌同样的表面上,而协调一致地行动,将细菌进往单一的方向前进。
一些细菌,如selenomonas ,鞭毛有组织外胞体。
③古菌鞭毛古菌的鞭毛表面上看起来类似细菌的鞭毛。但在20世纪90年代,研究人员曾发现了古菌和细菌鞭毛的具体不同,其中包括:
细菌鞭毛由流动的质子驱动,而古鞭毛几乎肯定由三磷酸腺苷驱动。
细菌细胞中往往有许多鞭毛的细丝,古菌鞭毛则由许多长丝组成一捆。

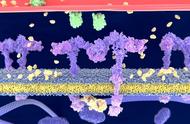
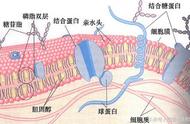
细胞膜,又称原生质膜或质膜(英语:cell membrane 或 plasma membrane 或 cytoplasmic membrane),为细胞结构中分隔细胞内、外不同介质和组成成分的界面。原生质膜普遍认为由磷脂质双层分子作为基本单位重复而成,即磷脂双分子层,其上镶嵌有各种类型的膜蛋白以及与膜蛋白结合的糖和糖脂。
原生质膜是细胞与周围环境和细胞与细胞间进行物质交换和信息传递的重要通道,可通过其上的孔隙和跨膜蛋白的某些性质,达到有选择性的、可调控的物质运输作用。
①学说1935年,J.F.Danielli和H.Davson提出双分子片层模型。1950年代,罗伯特森(J.D.Robertson)提出单位膜模型。1972年,美国加州大学的西摩·乔纳森·辛格(英语:Seymour Jonathan Singer)和加斯·L·尼科尔森(英语:Garth L. Nicolson)提出流动镶嵌模型(Fluid Mosaic Model)。
流动镶嵌模型是目前被最广泛接受和认可的观点。这种观点主张,构成膜的蛋白质和脂类分子具有镶嵌关系,而且膜的结构处于流动变化之中。许多实验结果支持流动镶嵌模型的模式。
②结构在流动镶嵌模型学说中,膜中的磷脂质分子以双层排列,构成了膜的网架,是膜的基质。磷脂质分子为双性分子,分为亲水头端和疏水尾端,双层磷脂质分子之头端皆朝向水相,疏水尾端则两两相接埋于膜内。而使脂双层分子之亲水头端的内层(面对细胞质之面)与外层(面对外界之面)之结构不对称原因,主要在于脂双层分子两亲水头端的化学组成不同。
膜的另一种主要成分是蛋白质,蛋白质分子有的嵌插在脂双层网架中,有的则粘附在脂双层的表面上。根据在膜上存在位置的不同,膜蛋白可分为两类,一是通过强疏水或亲水作用同膜脂牢固结合不易分开的,称为整合蛋白(Integral protein)或膜内在蛋白;二是附着在膜的表层,与膜结合比较疏松容易分离的,称为膜周边蛋白(Peripheral protein)或外在蛋白。
③功能原生质膜的功能主要有以下几个方面:
维持细胞的结构完整性,保护细胞内成分
细胞内外选择性物质运输的通道和桥梁
细胞抗原-抗体特异性识别的物质基础和位置
细胞表面绒毛、纤毛、鞭毛的着生位点
对于原核细胞而言,细胞质膜是很多催化生化反应的酶附着的位点。

细胞壁(英语:cell wall)是细胞的外层,在细胞膜的外面,细胞壁之厚薄常因组织、功能不同而异。它可以是坚韧的,有弹性,和有时坚硬的。它给细胞提供既有结构支承和保护,同时也作为一种过滤机制。植物、真菌(菌物)、藻类和原核生物都具有细胞壁,而支原体属细胞不具有细胞壁。
细胞壁的组成随着不同物种而变化,并可能取决于细胞的类型和发展阶段。陆生植物的初生细胞壁(primary cell wall)的组成是多糖类的纤维素,半纤维素和果胶。在细菌中,细胞壁的组成是肽聚糖。古菌细胞壁有各种组分物组成,并可能由糖蛋白的S层,假肽聚糖(英语:Pseudopeptidoglycan)或多糖组成的。真菌具有N-乙酰葡糖胺的聚合物甲壳素组成的细胞壁,和藻类通常具有糖蛋白和多糖组成的细胞壁。与众不同的是,硅藻具有一个由生物硅(英语:Biogenic silica)组成的细胞壁。其他辅助分子往往也锚定到细胞壁中,例如木质素和几丁质。
①成分与功能细胞壁是在细胞分裂、生长和分化过程中形成的,主要成分是纤维素和果胶,可用于支撑和维持植物细胞的形状。细胞壁分为三层,初生细胞壁(Primary cell wall)和次生细胞壁(Secondary cell wall)。细胞与细胞之间有胞间层(intercellular layer)分隔。所有植物细胞都有初生壁,其外面就是胞间层。次生壁在初生壁的里面,是在细胞停止生长后分泌形成的,可以增加细胞壁的厚度和强度,不易受到病原物多糖降解酶的直接攻击,但不是所有的细胞都具有次生壁。
次生壁又分内(S3)、中(S2)、外(S1)三层(在内层里面,有时还可出现一层),外层和内层都很薄,只有中层最厚,占次生壁厚度的70~90%,主要成分也是半纤维素、纤维素和木质素(lignin),极少含果胶,久之会开始进行不同程度的木质化,木聚糖逐渐分布于整个次生壁中,而木葡聚糖则局限分布于初生壁和胞间层,另外,角质(cutin)和木栓质(suberin)通常会埋入次生壁中。次生壁越厚,壁内的细胞腔就越小,等到细胞完全成熟后细胞腔呈现椭圆形。
②细菌细胞壁细胞膜外侧是细菌细胞壁。 细菌细胞壁由肽聚糖制成,其由多糖链制成,所述多糖链由含有D-氨基酸的不寻常肽交联。细菌细胞壁不同于分别由纤维素和甲壳素制成的植物和真菌的细胞壁。细菌的细胞壁也不同于不含肽聚糖的古菌细胞壁。尽管L型细菌可以在缺乏细胞壁的实验室中产生,但细胞壁对许多细菌的存活至关重要。抗生素青霉素能够通过阻止肽聚糖的交联来*死细菌,这会导致细胞壁变弱和溶解。 溶菌酶也可以破坏细菌细胞壁。
从广义上讲,细菌中有两种不同类型的细胞壁,革兰氏阴性和革兰氏阳性。这些名称源于细胞对革兰氏染色的反应,这是一种长期用于细菌种类分类的试验。
革兰氏阴性菌的细胞壁的结构和组成复杂,肽聚糖只有一薄层,在两层细胞膜之间的膜间腔中,而革兰氏阳性菌细胞壁厚得多,且只含有很多层肽聚糖。
③古菌细胞壁虽然不是真正独特,但古菌的细胞壁是不寻常的。尽管肽聚糖是所有细菌细胞壁的标准组分,但所有古菌细胞壁都缺乏肽聚糖,除了一组产甲烷菌(英语:Methanogen)外。 在那一组中,肽聚糖是一种与细菌中被发现的非常不同的一种改性的形式。古菌目前已知有四种类型的细胞壁。
一些古细菌的细胞壁由类似肽聚糖的假肽聚糖(英语:Pseudopeptidoglycan)构成,而另一些古菌既无肽聚糖,也无假肽聚糖,其细胞壁由多糖、糖蛋白或者蛋白构成。
【更多精彩文章,请关注微信公众号“世界民族与文明历史”】
,