梅塞尔森和凯恩斯等人证实了DNA的半保留复制机制。但是,关于DNA链延伸的具体机制,还存在一个“延伸方向悖论”,即所有DNA聚合酶均以5’-3’方向合成,而实验观察中两条反向平行的DNA链又都是连续延伸的。也就是说,下面的两个模型都缺乏实验支持。

DNA链延伸的两种模型。Proc Jpn Acad Ser B Phys Biol Sci. 2017;93(5):322-338.
解决这个问题的是名古屋大学的冈崎夫妇,冈崎令治(Reiji Okazaki)和冈崎恒子(Tsuneko Okazaki)。他们曾在科恩伯格实验室学习,回国后将复制方向问题作为自己的研究主题。冈崎认为,DNA复制过程中可能形成了很多较短的DNA片段,但用宏观方法难以分辨。比如放射自显影的每个颗粒对应的核酸片段其实大于1000个核苷酸,难以发现不连续的小片段。所以必须建立微观的、核苷酸水平的实验体系。
他们用放射性元素氚标记胸苷,在很短的时间内进行复制,以控制其仅在DNA链的末端掺入少数几个放射性核苷酸,即脉冲标记。经过计算,如果在37℃下合成,时间必须小于0.1秒才行,显然无法达成。所以他们采用了低温脉冲合成方法,在低温(20℃或更低)条件下将细菌暴露于氚标记的胸苷数秒,终于发现了长度为仅1,000-2,000个核苷酸的DNA片段(后来称为冈崎片段)。
当脉冲标记时间延长,放射性会从短的DNA片段转移到较长的DNA链。而对DNA连接酶的抑制会导致冈崎片段的积累。这些发现提示了DNA的不连续复制机制。1968年,冈崎令治应邀参加冷泉港研讨会,提出了DNA的不连续复制模型。

冈崎令治在冷泉港研讨会介绍不连续复制机制。Proc Jpn Acad Ser B Phys Biol Sci. 2017;93(5):322-338.
冈崎夫妇还发现冈崎片段合成需要RNA引物,以及前导链的连续复制等。可惜的是,冈崎令治在广岛读高中时遭遇原子弹辐射,1975年死于白血病,享年44岁。之后恒子继续研究,最终建立了半不连续复制(semi-discontinuous replication)模型。
在模型中,一条子代DNA链与复制叉同向,由5’向3’方向连续复制,称为前导链(leading strand);另一条链的延伸方向与复制叉相反,形成许多不连续的冈崎片段(Okazaki fragment),最后由DNA连接酶拼接成完整的DNA,称为滞后链(lagging strand)。

冈崎的半不连续复制模型。Proc Jpn Acad Ser B Phys Biol Sci. 2017;93(5):322-338.
图中模板DNA上的点表示引物RNA合成的起始信号序列。他们鉴定了模板DNA上的引物合成信号序列,并绘制在图中。结果表明,引物信号多于冈崎片段数。这说明在复制时可以选择不同的信号序列,具有一定的灵活性,也有助于复制不留空隙。
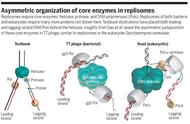
从半不连续模型上看,似乎前导链和滞后链是由两个酶分别合成,前进方向相反。但其实它们是由一个双核心的pol III复合物催化的,两条链前进的方向是相同的。这是布鲁斯·阿尔伯茨(Bruce Alberts)提出的 “长号” DNA循环模型(Trombone model)。