能量代谢的中心任务是生物体如何把外界环境中多种形式的最初能源转换成对一切生命活动都能使用的通用能源—— ATP 。对微生物来说,它们可利用的最初能源有三大类即:有机物、日光和还原态无机物。
一、异养微生物的生物氧化
生物氧化是发生在活细胞内的一系列产能性氧反应的总称。生物氧化的形式包括某物质与氧结合、脱氢或失去电子;生物氧化的过程可分为脱氢(或电子)、递氢(或电子)和受氢(或电子)三个阶段;生物氧化的功能则有产能、产还原力和产小分子中间代谢物三种。异养微生物氧化有机物的方式,根据氧化还原反应中电子受体的不同可分成发酵和呼吸两种类型,而呼吸以可分为有氧呼吸和无氧呼吸两种方式。
1 、发酵
发酵是指微生物细胞将有机物氧化释放的电子直接交给底物本身未完成氧化的某种中间产物,同时释放能量并产生各种不同的代谢产物。在发酵条件下有机化合物只是部分地被氧化,因此只释放出一小部分的能量。发酵过程的氧化是与有机物的还原偶联在一起的。被还原的有机物来自于初始发酵的分解代谢,即不需要外界提供电子受体。
发酵的种类有很多,可发酵的底物有糖类、有机酸、氨基酸等,其中以微生物发酵葡萄糖最为重要。生物体内葡萄糖被降解成丙酮酸的过程称为糖酵解,主要分为四种途径:EMP 、 HMP 、 ED 、磷酸解酮酶途径。
EMP 途径

整个 EMP 途径大致可分为两个阶段。第一阶段可认为是不涉及氧化还原反应及能量释放的准备阶段,只是生成两分子的主要中间代谢产物:甘油醛 -3- 磷酸。第二个阶段发生氧化还原反应,合成 ATP 并形成两分子的丙酮酸。在糖酵解过程中,有两分子 ATP 用于糖的磷酸化,但合成出四个分子的 ATP ,因此每氧化一个分子的葡萄糖净得两个 ATP 。
在两分子的 1 , 3- 二磷酯甘油酸的合成过程中,两分子 NAD 被还成为 NADH 。然而,细胞中的 NAD 供应是有限的,假如所有的 NAD 都转化为 NADH ,葡萄糖的氧化就得停止。因为甘油 -3- 磷酸的氧化反应只有在 NAD 存在时才能进行。这一路径可以通过将丙酮酸还原,使 NADH 氧化重新成为 NAD 而得以克服。例如在酵母细胞中丙酮酸被还原成为乙醇,并伴有 CO2 的释放。而在乳酸菌细胞中,丙酮酸被还原成乳酸。对于原核生物细胞,丙酮酸的还原途径是多样的,但有点是一致的:NADH 必须重新被还原成 NAD ,使得酵解过程中的产能反应得以进行。
EMP 途径可为微生物的生理活动提供 ATP 和 NADH ,其中间产物又可为微生物的合成代谢提供碳骨架,并在一定的条件下可逆转合成多糖。
HMP

HMP 途径是从葡萄糖 -6- 磷酸开始的, HMP 途径的一个循环的最终结果是一分子葡萄糖 -6- 磷酸转变成一分子甘油醛 -3- 磷酸,三分子 CO2 和六分子 NADPH 。一般认为 HMP 途径合成不是产能途径,而是为生物合成提供大量的还原力( NADPH )和中间代谢产物。如核酮糖 -5- 磷酸是合成核酸,某些辅酶及组氨酸的原料。另外 HMP 途径中产生的核酮糖 -5- 磷酸,还可以转化为核酮糖 -1 , 5- 二磷酸,在羧化酶作用下固定 CO2 ,对于光能自养菌、化通自养菌具有重要意义。
虽然这条途径中产生的 NADPH 可经呼吸链氧化产能, 1 摩尔葡萄糖经 HMP 途径最终可得到 35 摩尔 ATP ,但这不是代谢中的主要方式。因此,不能把 HMP 途径看作是产生 ATP 的有效机制。大多数好氧和兼性厌氧微生物中都有 HMP 途径,而且在同一微生物中往往同时存在 EMP 和 HMP 途径,单独具有 EMP 和 HMP 途径的微生物较少见。
ED 途径

ED 途径是在研究嗜糖假单胞菌时发现的,在 ED 途径中,葡萄糖 -6- 磷酸首先脱氢产生葡萄糖酸 -6- 磷酸,接着在脱水酶和醛缩酶的作用下,产生一个分子甘油醛 -3- 磷酸和一个分子丙酮酸。然后甘油醛 -3- 磷酸进入 EMP 途径转变成丙酮酸。一分子葡萄糖经 ED 途径最后生成两分子丙酮酸、一分子 ATP 、一分子 NADPH 和 NADH 。ED 途径在革兰代阴性菌中分布广泛,特别是假单胞菌和固氮的某些菌株较多存在。ED 途径可不依赖于 EMP 和 HMP 途径而单独存在,但对于靠底物水平磷酸化获得 ATP 的厌氧菌而言, ED 途径不如 EMP 途径。
磷酸解酮酶途径

磷酸解酮酶途径是明串珠菌在进行异型乳酸发酵过程中分解已糖和戊糖的途径。该途径的特征性酶是磷酸解酮酶,根据解酮酶的不同,把具有磷酸戊糖解酮酶的称为 PK 途径,把具有磷酸已糖解酮酶的称为 HK 途径。在糖酵解过程中生成的丙酮酸可被进一步代谢。在无氧条件下,不同的微生物分解丙酮酸后会积累不同的代谢产物。目前发现多种微生物可以发酵葡萄糖产生乙醇,能进行乙醇发酵的微生物包括酵母菌、根霉、曲霉和某些细菌。
根据在不同条件下代谢产物的不同,可将酵母菌利用葡萄糖进行的发酵分为三种类型:如果以乙醛(丙酮酸脱羧)为受体生成乙醇,这种发酵称为酵母的一型发酵;当环境中存在亚硫酸氢钠时,不能以乙醛作为受体,而以磷酸二羟丙酮作为受体时,产物为甘油,称为酵母的二型发酵;在弱碱性条件下( PH7.6 ),乙醛因得不到足够的氢而积累,两个乙醛分子间会发生歧化反应,一个作为还原剂形成乙酸,一个作为氧化剂形成乙醇,受体为磷酸二羟丙酮,发酵产物为甘油、乙醇和乙酸,称为酵母的三型发酵。
这种发酵方式不产生能量,只能在非生长的情况下进行。不同的细菌进行乙醇发酵时,其发酵途径也各不相同。如厌氧发酵单胞菌是利用 ED 途径分解葡萄糖为丙酮酸,最后得到乙醇。肠杆菌则是利用 EMP 途径来进行乙醇发酵。
许多细菌能利用葡萄糖产生乳酸,这类细菌称为乳酸细菌。根据产物的不同,乳酸发酵有三种类型:同型乳酸发酵(利用 EMP 途径产物只有乳酸)、异型乳酸发酵(利用 PK 乳酸及部分乙醇或乙酸)和双歧发酵(利用双歧双歧杆菌发酵葡萄糖产生乳酸的一条途径)。
2 、呼吸作用
微生物在降解底物的过程中,将释放出的电子交给 NAD ( P )、 FAD 或 FMN 等电子载体,再经电子传递系统传给外源电子受体,从而生成水或其他还原型产物并释放出能量的过程,称为呼吸作用。其中以分子氧作为最张终电子受体的称为有氧呼吸,以氧化型化合物作为最终电子受体的称为无氧呼吸。呼吸作用与发酵作用的根本区别在于:电子载体不是将电子直接传递给给底物降解的中间产物,而交给电子传递系统,逐步释放出能量后再将取终电子受体。
有氧呼吸
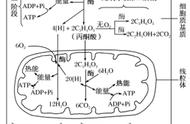
在发酵过程中,葡萄糖经过糖酵解作用形成的丙酮酸在厌氧化条件下转变成不同的发酵产物,而在有氧呼吸过程中,丙酮酸进入三羧酸循环( TCA )被彻底氧化成水和 CO2 ,同时释放出大量能量。
在 TCA 循环过程中,丙酮酸完全氧化为三个分子的 CO2 ,同时生成四分子的 NADH 和一分子 FADH 2 。NADH 和 FADH 2 可以电子传递系统重新被氧化,由此每一氧化一分子 NADH 可生成三个分子 ATP ,每氧化一分子 FADH 2 可生成两分子 ATP 。另外琥珀酰辅酶 A 在氧化成延胡索酸时,包含着底物水平磷酸化作用,由此产生一分子 GTP ,随后 GTP 转化 ATP 。因此每一次 TCA 循环可生成 15 分子 ATP 。
此外在糖酵解过程中产生的两分子 NADH 可经电子传递链系统重新被氧化,产生 6 分子 ATP 。在葡萄糖转变为两个分子丙酮酸时还可借底物水平磷酸化生成两分子 ATP 。因些需氧微生物在完全氧化葡萄糖的过程中总共可得到 38 分子的 ATP 。
在糖酵解和三羧酸循环过程中形成的 NADH 和 FADH 2 通过电子传递系统被氧化,最终形成 ATP 为微生物的生命活动提供能量。电子传递系统是由一系列氢和电子传递体组成的多酶氧化还原体系。NADH 、 FADH 2 以及其他还原型载体上的氢原子,以质子和电子的形式在其上进行定向传递;其组成酶系是定向有序的,又是不对称的地排列在原核微生物的细胞质膜上或是在真核微生物的线粒体内膜上。这些系统具有两种功能:一是从电子供体接受电子并将电子传递给电子受体;二是通过合成 ATP 把在电子传递过程中释放的一部分能量保存起来。电子传递系统中的氧化还原酶包括:NADH 脱氢酶、黄素蛋白、铁硫蛋白、细胞色素、醌及其化合物。
无氧呼吸
某些厌氧和兼性厌氧微生物在无氧条件下进行无氧呼吸。无氧呼吸的最终电子受体不是氧,而是像 NO3- 、 NO2- 、 SO42- 、 S2O32- 、 CO2 等这类外源受体。无氧呼吸也需要细胞色素等电子传递体。并在能量分级释放过程中伴随有磷酸化作用,也能产生较多的能量用于生命活动。但由于部分能量随电子转移给最终电子受体,所以生成的能量不如有氧呼吸产生的多。在无氧条件下,某些微生物在没有氧、氮或硫作为呼吸作用的最终电子受体时,可以磷酸盐代替,其结果生成磷化氢,一种易燃气体。在夜晚,气体燃烧会发出绿幽幽的光。
二、自养微生物的生物氧化和 CO2 的固定
一些微生物可以从氧化无机物获得能量,同化合成细胞物质,这类细菌称为化能自养微生物。它们在无机能源氧化过程中通过氧化磷酸化产生 ATP 。
(一)自养微生物的生物氧化
1 、氨的氧化
NH3 同亚硝酸 (NO2 - ) 是可以用作能源的最普通的无机氮化合物,能被硝化细菌所氧化,硝化细菌可分为两个亚群:亚硝化细菌和硝化细菌。氨氧化为硝酸的过程可分为两个阶段,先由亚硝化细菌将氨氧化为亚硝酸,再由硝化细菌将亚硝酸氧化为硝酸。由氨氧化为硝酸是通过这两类细菌依次进行的。硝化细菌都是一些专性好氧的革兰氏阳性细菌,以分子氧为最终电子受体,且大多数是专性无机营养型。它们的细胞都具有复杂的膜内褶结构,这有利于增加细胞的代谢能力。硝化细菌无芽抱,多数为二分裂殖,生长缓慢,平均代时在 l0h 以上,分布非常广泛。
2 、硫的氧化
硫杆菌能够利用一种或多种还原态或部分还原态的硫化合物 ( 包括硫化物、元素硫、硫代硫酸盐、多硫酸盐和亚硫酸盐 ) 作能源。H 2 S 首先被氧化成元素硫,随之被硫氧化酶和细胞色素系统氧化成亚硫酸盐,放出的电子在传递过程中可以偶联产生四个 ATP 。亚硫酸盐的氧化可分为两条途径,—是直接氧化成 SO42- 的途径,由亚硫酸盐 -- 细胞色素 c 还原酶和末端细胞色素系统催化,产生一个 ATP ;二是经磷酸腺苷硫酸的氧化的途径,每氧化一分子 SO42- 产生动 5 个 ATP 。
3 、铁的氧化
从亚铁到高铁状态的铁的氧化,对于少数细菌来说也是一种产能反应,但从这种氧化中只有少量的能量可以被利用。在低 pH 环境中这种菌能利用亚铁氧化时放出的能量生长。在该菌的呼吸链中发现了—种含铜蛋白质,它与几种细胞色素 c 和一种细胞色素 a 1 氧化酶构成电子传递链。在电子传递到氧的过程中细胞质内有质子消耗,从而驱动用 ATP 的合成。
4 、氢的氧化
氢细菌都是—些呈革兰氏阴性的兼性化能自养茵。它们能利用分子氢氧化产生的能量同化 CO 2 ,也能利用其他有机物生长。氢细菌的细胞膜上有泛醌、维生素 K 2 及细胞色素等呼吸链组分。在该菌中,电子直接从氢传递给电子传递系统,电子在呼吸链传递过程中产生了 ATP 。在多数氢细菌中有两种与氢的氧化有关的酶。—种是位于壁膜间隙或结合在细胞质膜上的不需 NAD 的颗粒状氧化酶,它能够催化以下反应:
H2 → 2H 十 2e -
该酶在氧化氢并通过电子传递系统传递电子的过程中,可驱动质子的跨膜运输,形成跨膜质子梯度为 ATP 的合成提供动力;另—种是可溶性氢化酶,它能催化氢的氧化,而使 NAD 还原的反应。所生成的 NADH 主要用于 CO 2 的还原。
(二) CO2的固定
CO2是自养微生物的唯一碳源,异养微生物也能利用CO2作为辅助的碳源。将空气中的CO2同化成细胞物质的过程,称为CO2的固定作用。微生物有两种同化CO2的方式,一类是自养式,另一类为异养式。在自养式中,CO2加在一个特殊的受体上,经过循环反应,使之合成糖并重新生成该受体。在异养式中, CO2被固定在某种有机酸上。因此异养微生物即使能同化 C02 ,最终却必须靠吸收有机碳化合物生存。
自养微生物同化CO2 所需要的能量来自光能或无机物氧化所得的化学能,固定 CO2的途径主要有以下三条:
1 、卡尔文循环 (Calvin cycle)
这个途径存在于所有化能自养微生物和大部分光合细菌中。经卡尔文循环同化 CO2的途径可划分为三个阶段见图:CO2的固定;被固定的CO2的还原;CO2 受体的再生。卡尔文循环每循环一次,可将六分子CO2同化成一分子葡萄糖,其总反应式为:
6CO2 18ATP 12NAD(P)H — C6H12O6 18ADP 12NAD(P) 18Pi
2 、还原性三竣酸循环固定CO2
这个途径见图是在光合细菌、绿琉细菌中发现的。还原羧酸环的第—步反应是将乙酰 CoA 还原羧化为丙酮酸,后者在丙酮酸羧化酶的催化下生成磷酸烯醇式丙酮酸,随即被羧化为草酰乙酸,草酰乙酸经一系列反应转化为琥珀酰 CoA ,再被还原羧化为 а - 酮戊二酸。а - 酮 戊二酸转化为柠檬酸后,裂解成乙酸和草酰乙酸。乙酸经乙酰 -CoA ,从而合成酶催化生成乙酰 CoA ,从而完成循环反应。每循环—次,可固定四分子CO2,合成一分子草酰乙酸,消耗三分子 ATP 、两分子 NAD(P)H 和—分子 FADH2 。
3 、还原的单羧酸环
这个体系与还原羧酸循环不同,不需要 ATP ,只要有 Fd(red) 就可运转。Fd(red) 由 H 2 或 NADH 2 提供电子生成。光合细菌也有可能利用这个体系把 CO2换成乙酸。
三、能量转换
在产能代谢过程中,微生物通过底物水平磷酸化和氧化磷酸化将某种物质氧化而释放的能量储存于 ATP 高能分子中,对光全微生物而言,则可通过光合磷酸化将光能转变为化学能储存于 ATP 中。
1 、底物水平磷酸化
物质在生物氧化过程中,常生成一些含有高能键的化合物,而这些化合物可直接偶联 ATP 或 GTP 的合成,这种产生 ATP 等高能分子的方式称为底物水平磷酸化。底物水平磷酸化既存在于发酵过程中,也存在于呼吸作用过程中。例如,在 EMP 途径中 1 , 3 —二磷酸甘油酸转变为 3 —磷酸甘油酸以及磷酸烯醇式丙酮酸转变为丙酮酸的过程中都分别偶联着—分子 ATP 的形成;在三磷酸循环过程中,琥珀酰辅酶 A 转变为琥珀酸时偶联着—分子 ATP 的形成。
2 、氧化磷酸化
物质在生物氧化过程中形成的 NADH 和 FADH 2 可通过位于线粒体内膜和细菌质膜上的电子传递系统将电子传递给氧或其他氧化型物质,在这个过程中偶联着 ATP 的合成,这种产生 ATP 的方式称为氧化磷酸化。一分子 NADH 和 FAD H 2 可分别产生 3 个和 2 个 ATP 。
3 、光合磷酸化
光合作用是自然界一个极其重要的生物学过程,其实质是通过光合磷酸化将光能转变成化学能,以用于从 CO 2 合成细胞物质。行光合作用的生物体除了绿色植物外,还包括光合微生物,如藻类、蓝细菌和光合细菌 ( 包括紫色细菌、绿色细菌、嗜盐菌等 ) 。它们利用光能维持生命,同时也为其他生物 ( 如动物和异养微生物 ) 提供了赖以生存的有机物。
(1) 光合色素
合色素是光合生物所特有的色素,是将光能转化为化学能的关键物质。共分三类:叶绿素 (chl) 或细菌叶绿素 (Bchl) ,类胡萝卜素和藻胆素。除光合细菌外,叶绿素 a 普遍存在于光合生物中,叶绿素 a 、 b 共同存在于高等植物、绿藻和蓝绿细菌中,叶绿素 c 存在于褐藻和硅藻中,叶绿素 d 存在于红藻中,叶绿素 e 存在于金黄藻中,褐藻和红藻也含有叶绿素 a 。细菌叶绿素具有和高等植物中的叶绿素相类似的化学结构,两者的区别在于侧链基团的不同,以及由此而导致的光吸收特性的差异。此外,叶绿素和细菌叶绿素的吸收光谱在不同的细胞中也有差异。
所有光合生物都有类胡萝卜素。类胡萝卜素虽然不直接参加光合反应,但它们有捕获光能的作用,能把吸收的光能高效地传给细菌叶绿素 ( 或叶绿素 ) 。而且这种光能同叶绿素 ( 或细菌叶绿素 ) 直接捕捉到的光能一样被用来进行光合磷酸化作用。此外胡萝卜素还有两个作用:一是可以作为叶绿素所催化的光氧化反应的猝灭剂,以保护光合机构不受光氧化损伤,二是可能在细胞能量代谢方面起辅助作用。
藻胆素因具有类似胆汁的颜色而得名,其化学结构与叶绿素相似,都含有四个吡咯环,但藻胆素没有长链植醇基,也没有镁原子,而且四个吡咯环是直链的。
(2) 光合单位
以往将在光合作用过程中还原一分子 C0 2 所需的叶绿素分子数称为光合单位。后来通过分析紫色细菌载色体的结构,获得了对光合单位的进一步认识。光合色素分布于两个“系统”,分别称为“光合系统 I ”和“光合系统 II ”。每个系统即为一个光合单位。这两个系统中的光合色素的成分和比例不同。一个光合单位由一个光捕获复合体和一个反应中心复合体组成。光捕获复合体含有菌绿素和类胡萝卜素,它们吸收一个光子后,引起波长最长的菌绿素 (P870) 激活,从而传给反应中心,激发态的 P870 可释放出一个高能电子。
(3) 光合磷酸化
光合磷酸化是指光能转变为化学能的过程。当—个叶绿素分子吸收光量子时,叶绿素性质上即被激活,导致叶绿素 ( 或细菌叶绿素 ) 释放一个电子而被氧化,释放出的电子在电子传递系统中的传递过程中逐步释放能量,这就是光合磷酸化的基本动力。
①环式光合磷酸化
光合细菌主要通过环式光合磷酸化作用产生 ATP ,这类细菌主要包括紫色硫细菌、绿色硫细菌、紫色非硫细菌和绿色非硫细菌。在光合细菌中,吸收光量子而被激活的细菌叶绿素释放出高能电子,于是这个细菌叶绿素分子即带有正电荷。所释放出来的高能电子顺序通过铁氧还蛋白、辅酶 Q 、细胞色素 b 和 c ,再返回到带正电荷的细菌叶绿素分子。在辅酶 Q 将电子传递给细胞色素 c 的过程中,造成了质子的跨膜移动,为 ATP 的合成提供了能量见图。在这个电子循环传递过程中,光能转变为化学能,故称环式光合磷酸化。环式光合磷酸化可在厌氧条件下进行,产物只有 ATP ,无 NADP(H) ,也不产生分子氧。
②非环式光合磷酸化
高等植物和蓝细菌与光合细菌不同,它们可以裂解水,以提供细胞合成的还原能力。它们含有两种类型的反应中心,连同天线色素、初级电子受体和供体一起构成了光合系统 I 和光合系统 II ,这两个系统偶联,进行非环式光合磷酸化见图。在光合系统 I 中,叶绿素分子 P 700 吸收光子后被激活,释放出—个高能电子。这个高能电子传递给铁氧还蛋白 (Fd) ,并使之被还原。还原的铁氧还蛋白在 Fd :NADP 还原酶的作用下,将 NADP 还原为 NADPH 。用以还原 P 700 的电子来源于光合系统 II 。在光合系统 II 中,叶绿素分子 P 680 吸收光子后,释放出一个高能电子。后者先传递给辅酶 Q ,再传给光合系统 I ,使 P 700 还原。失去电子的 P 680 ,靠水的光解产生的电子来补充。高能电子从辅酶 Q 到光合系统 I 的过程中,可推动 ATP 的合成。非环式光合磷酸化的反应式为:
2NADP 2ADP 2Pi 2H2O → 2NADPH 2H 2ATP O2
有些光合细菌虽然只有一个光合系统,但也以非环式光合磷酸化的方式合成 ATP ,如绿硫细菌和绿色细菌。从光反应中心释放出的高能电子经铁硫蛋白、铁氧还蛋白、黄素蛋白,最后用于还原 NAD 生成 NADH 。反应中心的还原依靠外源电子供体,如 S 2- 、 S 2 O 3 2- 等。外源电子供体在氧化过程中放出电子,经电子传递系统传给失去了电子的光合色素,使其还原,同时偶联 ATP 的生成。由于这个电了传递途径也没有形成环式,故也称为非环式光合磷酸化。
,