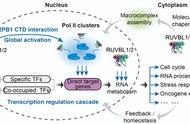
抢帽过程中的构象变化。Trends Microbiol. 2019 May; 27(5): 398–407.
RdRP容易出错,所以RNA病毒平均每轮复制产生一个突变。这使病毒能够快速适应新的生存环境,如更换宿主、逃避宿主免疫系统、产生抗药性等。所以流感病毒等每年都会产生新的突变株,大约每十年会由于出现一种重大突变而引起一次大流行。
现已发现,RdRP并非仅存在于病毒中。RdRP在植物和线虫中都有发现,有报道称果蝇的Elp1是一种新型的RdRP(Proc Natl Acad Sci USA. 2009)。人类端粒酶催化亚基(hTERT)可以与线粒体核糖核酸内切酶(RMRP)形成一种核糖核蛋白复合物,具有RdRP活性(Nature. 2009)。
以上研究表明,RdRP在真核生物中可能也有重要作用,例如调控非编码RNA等。现在较为确定的是RdRP可以参与SiRNA介导的转录后基因沉默(PTGS)。

RdRP以两种方式催化SiRNA形成。Cancer Sci. 2015 Nov; 106(11): 1486–1492.
RdRP可以通过Dicer依赖和非依赖的两种途径产生SiRNA。前者是RdRP先合成较长的双链RNA,再被Dicer加工成SiRNA;后者是RdRP直接从头合成SiRNA而无需酶切。
所以TERT可能具有双重聚合酶活性,端粒酶活性可以保持端粒结构并有助于细胞永生化,而RdRP活性则介导RNA合成和异染色质维持,并调节有丝分裂进程和肿瘤干细胞性状。也就是说,TERT作为抗肿瘤治疗的分子靶标可以通过两种机制起作用(Cancer Sci. 2015)。

TERT可以作为抗癌靶标。Cancer Sci. 2015 Nov; 106(11): 1486–1492.
参考文献:
1. Ng KK, et al. Structure-Function Relationships Among RNA-Dependent RNA Polymerases. Curr Top Microbiol Immunol. 2008; 320: 137–156.
2. Shu B, Gong P. The uncoupling of catalysis and translocation in the viral RNA-dependent RNA polymerase. RNA Biol. 2017 Oct 3;14(10):1314-1319.
3. Wu J, Gong P. Visualizing the Nucleotide Addition Cycle of Viral RNA-Dependent RNA Polymerase. Viruses. 2018 Jan; 10(1): 24.
4. Chetverin AB. Thirty Years of Studies of Qβ Replicase: What Have We Learned and What Is Yet to Be Learned? Biochemistry (Mosc). 2018 Jan;83(Suppl 1):S19-S32.
5. Samatov TR, et al. Expressible molecular colonies. Nucleic Acids Res. 2005; 33(17): e145.
6. Walker AP, Fodor E. Interplay between Influenza Virus and the Host RNA Polymerase II Transcriptional Machinery. Trends Microbiol. 2019 May; 27(5): 398–407.
7. Lipardi C, Paterson BM. Identification of an RNA-dependent RNA polymerase in Drosophila involved in RNAi and transposon suppression. Proc Natl Acad Sci USA. 2009;106:15645–15650.
8. Maida Y, Yasukawa M, Furuuchi M et al An RNA‐dependent RNA polymerase formed by TERT and the RMRP RNA. Nature 2009; 461: 230–5.
9. Maida Y, Masutomi K. Telomerase reverse transcriptase moonlights: Therapeutic targets beyond telomerase. Cancer Sci. 2015 Nov; 106(11): 1486–1492.
,