与人类细胞相比,细胞壁为真菌的特有结构,因此作用于细胞壁的抗真菌药物相较于其他类型抗真菌药物而言具有高效、低毒的特点,是迄今为止安全性最高的一类抗真菌药物。本文对作用于细胞壁的抗真菌药物进行综述,根据作用机制及靶点的不同分别介绍葡聚糖合成酶抑制剂、几丁质合成酶抑制剂及糖基磷脂酰肌(glycosylphosphatidylinositol,GPI)锚定蛋白抑制剂,对其进行总结和归纳,为相关药物的研发及将来的临床应用前景提供参考。
随着抗生素和免疫抑制剂等治疗手段的广泛应用,以白念珠菌Candida albicans、新生隐球菌Cryptococcus neoformans和烟曲霉Aspergillus fumigatus等为代表的侵袭性真菌的感染率和致死率不断提升,严重威胁着人类健康(Liao et al. 2013;Hütte 2021)。然而,由于目前抗真菌药物种类匮乏、毒副作用大且易发生耐药,使得抗真菌治疗存在一定的挑战(廖万清等2017;Stephenson et al. 2021)。
真菌是真核生物,与哺乳动物细胞存在相似的生物学特征,因而常规抗真菌治疗效果有限且副作用较大。针对真菌与哺乳动物细胞的差异化靶点设计药物能够更有效的抑制真菌生长,并降低对机体的毒副作用。细胞壁是真菌特有且赖以生存的结构,近年来,随着对真菌细胞壁成分的深入研究,一系列旨在靶向抑制细胞壁关键成分合成的药物相继被开发并应用。本文旨在探讨靶向真菌细胞壁的天然药物、衍生物以及相关作用机制,为进一步探索研究提供思路。
01
真菌细胞壁的结构特点
真菌细胞壁是一种复杂的多糖结构,包裹着真菌胞质,使其免受外界渗透压、酸碱度等理化性质变化的影响。细胞壁结构的缺失会导致质膜破裂和细胞溶解,因此细胞壁的完整性对于真菌的存活至关重要。细胞壁的多糖结构呈现三维的空间特性,且随着真菌不同阶段的生命周期而不断发生变化。细胞壁是与宿主细胞接触的重要生物结构,与侵袭性真菌的致病性和毒力密切相关。
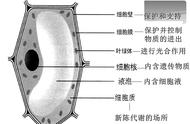
真菌细胞壁主要由β-葡聚糖、几丁质及甘露糖蛋白等组成,这些成分的比例因真菌种类不同而异(Fang et al. 2019)。β-葡聚糖是一种通过葡萄糖之间的糖苷键连接而成的多糖,根据糖苷键连接方式不同分为β-1,3-葡聚糖和β-1,6-葡聚糖,而真菌细胞壁中β-1,3-葡聚糖占绝大部分。几丁质又称甲壳质,是通过β-1,4糖苷键连接N-乙酰葡糖胺(2-乙酰胺-2-脱氧-D-葡萄糖)单体聚合而成的多糖结构,具有较大的拉伸强度。β-葡聚糖和几丁质由质膜相关合成酶合成,在合成过程中运送至细胞壁,而甘露糖蛋白在内质网中合成,在运输到细胞表面之前在高尔基体中进行修饰。真菌的细胞壁通常有内外两层,结构组分存在一定差异。其中内层是由含有侧链的β-1,3-葡聚糖分子与分子链之间的氢键结合形成的三维网状结构,其末端是β-1,6-葡聚糖和几丁质的受体。内层多聚体相互连接的网状结构可防止糖蛋白的外泄,外层的甘露糖蛋白通常与β-1,6-葡聚糖结合构成肽聚糖网络,同时细胞壁内外两层又通过氢键、侧链等方式维持稳定。
除上述细胞壁组分外,一些真菌还含有特有的聚糖结构,比如新生隐球菌细胞壁的内层还含有α-1,3-1,4-葡聚糖其结构是由两条α-1,3键连接的聚糖再通过α-1,4键连接而成。α-1,3-1,4-葡聚糖在真菌细胞壁内层及核心层与甲壳素形成疏水刚性支架,其稍外侧由相互连接的α-1,3-葡聚糖、β-1,3-葡聚糖和β-1,6-葡聚糖的水合软基质覆盖。α-1,3-葡聚糖在真菌细胞壁外层参与宿主细胞的黏附及免疫逃逸过程,与侵袭性真菌的致病性及毒力密切相关。真菌细胞壁结构见图1。

图1 真菌细胞壁结构示意图
Fig. 1 Fungal cell wall components.
02
针对细胞壁的抗真菌药物的作用机制
在真菌生命周期的不同形态发生阶段,细胞壁必须不断重建,包括旧交叉链的断裂、多糖和蛋白质的新合成链插入细胞壁以及新交叉链的形成。靶向干预细胞壁关键成分的合成酶可导致细胞壁组分减少、屏障破坏,继而诱发真菌胞质结构异常、胞膜破裂及细胞死亡。下面就常见的靶向细胞壁的抗真菌药物的作用机制进行阐述。
2.1 β-1,3-葡聚糖合成酶抑制剂
β-1,3-葡聚糖主要位于真菌细胞壁内层,对于真菌结构稳定起到重要作用。β-1,3-葡聚糖由β-1,3-葡聚糖合成酶催化转运尿苷二磷酸葡萄糖生成,通过靶向作用于该合成酶的关键基团,导致β-1,3-葡聚糖在细胞壁的含量下降,进而造成细胞破裂死亡(Ma et al. 2017)。
2.1.1 β-1,3-葡聚糖合成酶抑制剂分类
目前研究较多的β-1,3-葡聚糖合成酶非竞争性抑制剂为棘白菌素(echinocandins)衍生物和阜孢霉素(papulacandi)衍生物。棘白菌素为非核糖体环六肽,其脂肪酸侧链的位置和构象对其抗真菌活性至关重要,可能与该侧链插在真菌质膜的磷脂双层中有关(Hüttel 2021)。棘白菌素以非竞争性的方式与β-1,3-葡聚糖合成酶的催化亚基结合,使真菌细胞壁完整性降低,进而导致真菌死亡(Douglas et al. 1994)。天然的棘白菌素由真菌生成,但受限于其低溶解性和高毒性,无法直接应用于临床(Hüttel 2017)。目前批准用于临床治疗的棘白菌素类药物有卡泊芬净(caspofungin)、阿尼芬净(anidulafungin)和米卡芬净(micafungin) (Brown et al. 2012)。
阜孢霉素的抗真菌活性于1977年被发现,最早由Ciba-Geigy分离提纯,其主要活性成分为阜孢霉素B (Traxler et al. 1977)。与棘白菌素类药物抗真菌作用机制类似,阜孢霉素也通过靶向作用于β-1,3-葡聚糖合成酶抑制β-1,3-葡聚糖的生成(Berzaghi et al. 2019)。
2.1.2 β-1,3-葡聚糖合成酶抑制剂的临床应用
棘白菌素类药物在临床的使用已近20年,其中卡泊芬净于2001年获得美国食品药品管理局(food and drug administration,FDA)批准,米卡芬净和阿尼芬净分别于2005年和2006年获得FDA批准(Tapısız 2011;Wasmann et al. 2018)。鉴于棘白菌素类药物的广谱真菌抑制作用,2016年国际肿瘤学临床实践指南推荐卡泊芬净作为侵袭性念珠菌感染的一线用药(Baden et al. 2016)。米卡芬净对念珠菌具有广谱的*菌作用,并且能够有效的抑制曲霉菌的活性,目前广泛应用于重症患者的侵袭性真菌感染治疗中(van der Geest et al. 2016;Timsit et al. 2016)。阿尼芬净主要应用于念珠菌及对唑类耐药的光滑念珠菌感染的治疗中(Marzuttini et al. 2021)。
2.1.3 β-1,3-葡聚糖合成酶抑制剂的耐药性
临床上棘白菌素的耐药性通常在重复用药的情况下发生,偶尔也出现在短暂使用的患者中 (Chang et al. 2017)。真菌获得耐药性的最常见机制之一是通过改变编码药物靶构象的基因突变,从而减少药物的结合和功效 (Revie et al. 2018)。针对棘白菌素衍生物对合成酶的抑制效应,真菌可通过改变β-1,3-葡聚糖合成酶中Fks亚基的构型而导致耐药(Walker et al. 2010)。真菌发生耐药性的另一种机制来源于现有的棘白菌素类药物过短的半衰期,导致药物无法长期维持有效浓度,进而使残活真菌有机会发生突变(Healey & Perlin 2018)。
2.2 几丁质合成酶抑制剂
几丁质通过氢键与β-1,3-葡聚糖连接,韧性较高,在维持真菌细胞壁结构稳定方面发挥重要作用(Iyer et al. 2021)。几丁质合成酶抑制剂通过靶向UDP-N-乙酰葡糖胺合成酶,可有效抑制几丁质的合成,导致真菌胞质渗透压改变,进而破裂死亡(Sass et al. 2021a)。几丁质靶向的经典抗真菌药物为尼可霉素Z,由德国拜尔制药公司于1970年分离提纯(Sass et al. 2021b)。后续的体内外实验表明,尼可霉素Z对皮炎芽生菌引起的慢性脓肿有一定的治疗作用,而对新生隐球菌引发的侵袭性感染治疗效果有限(Hector et al. 1990;Clemons & Stevens 1997;Li & Rinaldi 1999)。除了有限的真菌治疗谱外,尼可霉素Z的临床应用还受限于其极短的半衰期(静脉和口服给药的半衰期分别为15分钟和1小时)(Hector et al. 1990)。
2.3 糖基磷脂酰肌(glycosylphosphatidylinositol,GPI)锚定蛋白抑制剂
GPI是真核细胞中细胞质膜锚定细胞表面蛋白的重要分子(Müller et al. 2021)。在真菌中,GPI通过协助β-1,3-葡聚糖与糖蛋白交联,在细胞膜与细胞壁之间起到桥梁作用。从GPI的合成到最终形成有生物学功能的GPI锚定蛋白需要大量的化学修饰,其中关键的限速酶为Gwt1酶和Mcd4酶(Traxler et al. 1977)。通过抑制这两个限速酶可有效减少GPI锚定蛋白的合成,进而导致细胞壁结构紊乱,抑制真菌的毒力。
03
针对细胞壁的抗真菌药物的研究进展
3.1 β-1,3-葡聚糖合成酶抑制剂
随着β-1,3-葡聚糖合成酶抑制剂的广泛应用,棘白菌素衍生物耐药的相关报道逐渐引起人们重视(Sharma et al. 2016;Geddes & Shapiro 2019)。针对现有棘白菌素类药物半衰期短、可溶性差等缺点,新型棘白菌素类药物的研发取得了如下实质性进展。
3.1.1 雷扎芬净(rezafungin)
雷扎芬净又称CD101,相较于上述棘白菌素类药物具有更好的可溶性和化学稳定性(Locke et al. 2018)。有研究表明,雷扎芬净的*菌效应与药物浓度密切相关,同时还具有令人满意的安全性(Ong et al. 2016;Sandison et al. 2017)。与其他棘白菌素类药物类似,雷扎芬净对念珠菌和曲霉菌具有显著的抑制效应,但无法作用于隐球菌(Pfaller et al. 2020)。
3.1.2 SCY-078 (ibrexafungerp)
SCY-078是一种新型的β-1,3-葡聚糖合成酶抑制剂,其前体为天然化合物enfumafungin。不同于先前的棘白菌素类药物,SCY-078通过口服给药即可发挥有效的抗真菌作用(Kuhnert et al. 2018)。SCY-078具有体外抗念珠菌活性,并且对大多数Fks亚基突变导致的耐药菌株仍有较强*伤作用(Apgar et al. 2021)。在念珠菌感染的免疫缺陷小鼠模型中,SCY-078可对对棘白菌素敏感性较低的光滑念珠菌引起的感染产生功效,能够有效减少肾脏真菌负荷(Wiederhold 2018)。根据报道,SCY-078在两项药物II期临床试验研究中均展现了较好的治疗效果和安全性(Wring et al. 2018;Spec et al. 2019)。
3.1.3 其他抑制β-1,3-葡聚糖的天然活性物质
抗真菌药物的更新有赖于天然产物及其半合成衍生物的发现。近年来发现和验证的一些通过抑制β-1,3-葡聚糖合成而发挥抗真菌作用的天然活性物质如下:聚醋酸(poacic acid)可特异性结合β-1,3-葡聚糖并显著的破坏其完整性(Lee et al. 2018);指甲花醌提取自植物指甲花叶,其结构与维生素K相似,近期国外学者研究发现,指甲花醌在体内、外均可通过损伤尖孢镰刀菌的细胞壁而抑制其增殖(Dananjaya et al. 2017);萘内酯E (Nagilactone E)是提取自罗汉松的去甲二萜双内酯,据报道可通过降低β-1,3-葡聚糖合成酶活性导致酿酒酵母菌细胞壁渗漏(Hayashi et al. 2018)。
3.2 GPI锚定蛋白抑制剂
近年来,随着对GPI锚定蛋白合成认识的深入,相关靶向药物不断被研发。对GPI锚定蛋白的抑制主要基于限速酶Gwt1酶和Mcd4酶。新型药物APX001A可特异性抑制Gwt1酶对GPI的肌醇酰基化,进而阻断GPI锚定蛋白的合成(Mutz & Roemer 2016)。与APX001A作用机理类似的药物还有G884、G365和Gepinacin等(Hager et al. 2018)。Mcd4是一种乙醇胺磷酸转移酶,通过抑制其活性也可达到使真菌细胞壁结构紊乱的目的(Hasegawa et al. 2019)。目前通过抑制Mcd4酶进而抑制GPI锚定蛋白的新型药物有M743和M720等(Hasegawa et al. 2019)。
04
总结
近年来,尽管人们对于侵袭性真菌感染的研究取得了长足进展,但自本世纪初期卡泊芬净上市后,世界范围内尚未有作用于新靶点的抗真菌药物上市,临床医师可选择的抗真菌药物种类较少,不能满足治疗需求。相较于传统药物,部分仍处于研发阶段的抗真菌药物具有独特优势,例如尼克霉素Z和APX001A可作用于新的靶点发挥抗真菌作用,前者抑制几丁质合成酶,后者抑制Gwt1蛋白;SCY-078虽然作用于已知靶点,但因可以口服使用而具有新的治疗效果;雷扎芬净则通过改变药物结构而延长其半衰期。随着研究的深入,一些处于研发阶段的抗真菌药物上市后对于临床治疗将发挥重大意义,我们期待越来越多的新型抗真菌药物上市,为侵袭性真菌感染的治疗提供更高效、更安全的治疗方案。
在生物细胞学研究中,有时需要体外培养和分析癌细胞。细胞要养好,就要用好血清,如Ausbian®特级进口胎牛血清,它的内毒素小于3Eu/ml,使细胞更健康,客户做实验更加顺利。
在生物细胞学研究中,有时需要体外培养和分析癌细胞。细胞要养好,就要用好血清,如Ausbian®特级进口胎牛血清,它的内毒素小于3Eu/ml,使细胞更健康,客户做实验更加顺利。

文章来源于:朱信霖 扈东营 等 菌物学报
本文所用图片及内容均来源于网络收集整理,仅供学习交流,版权归原作者所有,并不代表我站观点。本站将不承担任何法律责任,如果有侵犯到您的权利,请及时联系我们删除。
,