糖类分解代谢的第二阶段,是糖酵解。在生化中,酵解是指酶催化葡萄糖降解成丙酮酸并生成ATP和NADH的过程。它是各种生物体中葡萄糖分解产生能量的共同代谢途径。
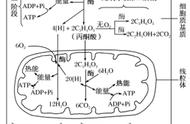
丙酮酸是代谢网络中的重要节点之一,联系着酵解、糖异生、三羧酸循环和氨基酸代谢。如果需要继续氧化分解,丙酮酸就会进入线粒体,经三羧酸循环彻底氧化生成CO2和水,酵解生成的NADH则经呼吸链氧化产生ATP和水。这就是分解代谢的第三阶段了。

丙酮酸联系着多条代谢途径
有一个概念容易与酵解混淆,就是“发酵”。微生物中的发酵大致是指微生物培养,而生化中的发酵是指一种特殊的葡萄糖或有机物降解产生ATP的过程,其中有机物既是电子供体,又是电子受体。这就是说,要把中间产生的NADH再氧化成NAD 。不同生物采用的具体反应不同,人体是用丙酮酸氧化NADH,生成乳酸;微生物中可以产生多种多样的产物,如乙醇、乙酸、丙酸、丙酮、丁醇、丁酸、琥珀酸、丁二醇等。
标准的酵解过程共有10步,前5步是准备阶段,葡萄糖分解为三碳糖,消耗2分子ATP;后5步是放能阶段,三碳糖生成丙酮酸,共产生4分子ATP。总过程需10种酶,都在细胞质中,多数需要Mg2 。酵解过程中所有的中间物都是磷酸化的,可防止从细胞膜漏出、保存能量,并有利于与酶结合。酵解的具体反应过程各种教材均有讲解,这里只介绍一些容易忽略的内容。

糖酵解过程
酵解的第一步由己糖激酶(hexokinase,HK,EC 2.7.1.1)催化,可作用于D-葡萄糖、果糖和甘露糖。人体己糖激酶有四种同工酶, 其中HK4对葡萄糖的特异性强,又称葡萄糖激酶(glucokinase,GK)。GK主要分布在肝脏,Km较高(10mM),不受产物6-磷酸葡萄糖的别构抑制,主要用于调控餐后血糖。而其它同工酶用于为肌肉、脑等组织提供能量。详见《己糖激酶的同工酶及其对糖酵解的调控作用》一文。

酵母己糖激酶的诱导契合,引自PDB-101
酵解的第二步反应是可逆的,反应方向由两侧的相对浓度控制。磷酸葡萄糖异构酶受磷酸戊糖支路的中间物(如6-磷酸葡萄糖酸)竞争性抑制。戊糖支路通过这种方式抑制酵解和有氧氧化,pH降低使抑制加强,减少酵解,以免组织过酸。
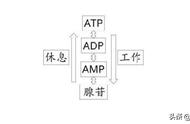
催化第三步反应的磷酸果糖激酶(6-phosphofructokinase,PFK,EC2.7.1.11)是酵解的限速酶,所以调节物很多。ATP、柠檬酸、磷酸肌酸、脂肪酸、DPG是负调节物;1,6-二磷酸果糖、2,6-二磷酸果糖、AMP、ADP、磷酸、环AMP等是正调节物。
6-磷酸果糖还可以被磷酸果糖激酶-2(PFK-2,EC2.7.1.105)催化,产生2,6-二磷酸果糖。它是PFK最强的别构激活剂,在酵解和异生的协调中起着重要作用。

PFK的变构,引自PDB-101
PFK有三种同工酶,A在心肌和骨骼肌中,对磷酸肌酸、柠檬酸和磷酸敏感;B在肝和红细胞中,对DPG敏感;C在脑中,对ATP和磷酸敏感。这种敏感性的差异,不仅因为各种组织中浓度不同,更重要的是其浓度变化幅度不同。例如,大鼠在运动和休息时ATP含量仅差0.8 ug/g肌肉,不能显著改变PFK活力;而磷酸肌酸含量是ATP的3-4倍,并且在肌肉收缩开始后几秒,即从20 mM下降到10~5 mM,所以会显著增加PFK活力。
第五步异构化完成之后,一分子葡萄糖转变为2分子3-磷酸甘油醛,消耗了2分子ATP。需要注意的是,原来葡萄糖的3,2,1位和4,5,6位变成了甘油醛的1,2,3位。这一点在分析碳原子去向的题目中会用到。
,