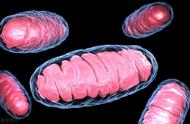
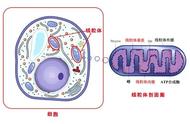
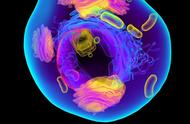
线粒体(mitochondrion)是一种存在于大多数细胞中的由两层膜包被的细胞器,是细胞中制造能量的结构,是细胞进行有氧呼吸的主要场所,被称为"power house"。其直径在0.5到1.0微米左右。
除了溶组织内阿米巴、篮氏贾第鞭毛虫以及几种微孢子虫外,大多数真核细胞或多或少都拥有线粒体,但它们各自拥有的线粒体在大小、数量及外观等方面上都有所不同。
线粒体拥有自身的遗传物质和遗传体系,但其基因组大小有限,是一种半自主细胞器。除了为细胞供能外,线粒体还参与诸如细胞分化、细胞信息传递和细胞凋亡等过程,并拥有调控细胞生长和细胞周期的能力。
主要功能:1,能量转化
线粒体是真核生物进行氧化代谢的部位,是糖类、脂肪和氨基酸最终氧化释放能量的场所。线粒体负责的最终氧化的共同途径是三羧酸循环与氧化磷酸化,分别对应有氧呼吸的第二、三阶段。
2,三羧酸循环
糖酵解中生成的每分子丙酮酸会被主动运输转运穿过线粒体膜。进入线粒体基质后,丙酮酸会被氧化,并与辅酶A结合生成CO2、还原型辅酶Ⅰ和乙酰辅酶A。
乙酰辅酶A是三羧酸循环(也称为“柠檬酸循环”或“Krebs循环”)的初级底物。参与该循环的酶除位于线粒体内膜的琥珀酸脱氢酶外都游离于线粒体基质中。
在三羧酸循环中,每分子乙酰辅酶A被氧化的同时会产生起始电子传递链的还原型辅因子(包括3分子NADH和1分子FADH2)以及1分子三磷酸鸟苷(GTP)。
3,氧化磷酸化
NADH和FADH2等具有还原性的分子(在细胞质基质中的还原当量可从由逆向转运蛋白构成的苹果酸-天冬氨酸穿梭系统或通过磷酸甘油穿梭作用进入电子传递链)在电子传递链里面经过几步反应最终将氧气还原并释放能量,其中一部分能量用于生成ATP,其余则作为热能散失。
在线粒体内膜上的酶复合物(NADH-泛醌还原酶、泛醌-细胞色素c还原酶、细胞色素c氧化酶)利用过程中释放的能量将质子逆浓度梯度泵入线粒体膜间隙。
虽然这一过程是高效的,但仍有少量电子会过早地还原氧气,形成超氧化物等活性氧(ROS),这些物质能引起氧化应激反应使线粒体性能发生衰退。
当质子被泵入线粒体膜间隙后,线粒体内膜两侧便建立起了电化学梯度,质子就会有顺浓度梯度扩散的趋势。质子唯一的扩散通道是ATP合酶(呼吸链复合物V)。
当质子通过复合物从膜间隙回到线粒体基质时,电势能被ATP合酶用于将ADP和磷酸合成ATP。这个过程被称为“化学渗透”,是一种协助扩散。
彼得·米切尔就因为提出了这一假说而获得了1978年诺贝尔奖。1997年诺贝尔奖获得者保罗·博耶和约翰·瓦克阐明了ATP合酶的机制。
4,储存钙离子
线粒体可以储存钙离子,可以和内质网、细胞外基质等结构协同作用,从而控制细胞中的钙离子浓度的动态平衡。线粒体迅速吸收钙离子的能力使其成为细胞中钙离子的缓冲区。
在线粒体内膜膜电位的驱动下,钙离子可由存在于线粒体内膜中的单向运送体输送进入线粒体基质;排出线粒体基质时则需要钠-钙交换蛋白的辅助或通过钙诱导钙释放(calcium-induced-calcium-release,CICR)机制。
在钙离子释放时会引起伴随着较大膜电位变化的“钙波”(calcium wave),能激活某些第二信使系统蛋白,协调诸如突触中神经递质的释放及内分泌细胞中激素的分泌。线粒体也参与细胞凋亡时的钙离子信号转导。