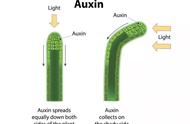
3、生长素运输与生长素差异分布
在单个细胞水平上,生长素的差异积累是通过核生长素信号通路感知和发挥作用的,该通路调节基因的表达甚至改写细胞命运(Keithanne Mockaitis and Mark Estelle, 2008)。生长素的差异分布对许多发育过程至关重要,因此,调控生长素分布或细胞生长素信号会强烈干扰相关的发育过程(Friml et al., 2003;Didier Reinhardt, 2005;Heisler et al., 2005;Sabatini et al., 1999;Friml et al., 2004)。那么一个重要的问题来了,生长素分布的差异是如何产生和维持的?生长素极性运输和局部生物合成都将导致生长素分布的差异。值得注意的是,不同的内部和外部信号已被证明可以调节生长素的生物合成(在关键酶的转录水平)和极性运输(在转录水平、细胞表面丰度和运输组分的极性定位)(Steffen Vanneste and Jirí Friml, 2009)。因此,运输依赖的生长素差异分布代表了生长素信号通路中额外水平的调控和信号整合。
3.1、化学渗透假说的模型
解释生长素极性运输机制的尝试最初依赖于生化和生理研究,这些研究使得在20世纪70年代形成了化学渗透模型(Rubery P H and Sheldrake A R, 1974;Raven J A, 1975)。生长素IAA是一种弱酸,它主要依赖于转运体的跨细胞途径进行转运,该途径主要是利用了质膜H -ATP酶产生的细胞外空间和细胞质之间的pH差异。质外体的pH值为5.5,其中IAA部分质子化,可以通过质膜被动扩散进入细胞。这种被动内流是由生长素吸收载体辅助进行的。此外,在更基本的细胞质环境中,IAA以带电的、不透细胞膜的形式存在,并被困在细胞内,因此它运输出细胞需要生长素外排载体(图1)。利用放射性标记的生长素衍生物进行运输实验,结果表明生长素的运输是定向的(极性的),主要的运输路线为从茎尖流向根。为了从机制上解释这种方向性,化学动力学模型假设外排载体的不对称(极性)质膜定位将决定生长素在细胞间的流动方向(Rubery P H and Sheldrake A R, 1974;Raven J A, 1975)。这一理论认为生长素载体在单细胞水平上的极性和极性靶向决定了细胞间信号的定向传输,从而介导了整个组织的发育。因此,外排载体的极性亚细胞定位对生长素的流动方向至关重要!
3.2、生长素运输蛋白质
化学渗透假说最初假设生长素内流和外排载体的存在,是在缺乏其分子特性信息的情况下确定的。后来,得益于拟南芥作为植物分子遗传学模式生物的成功建立,生长素转运体的身份才逐渐被揭示。这些包括(i)AUX1/ LAX内流蛋白,它介导生长素进入细胞;(ii)ABCB/PGP蛋白,介导ATP依赖的生长素通过质膜转运;(iii)PIN外排蛋白,通过极性亚细胞定位决定生长素运输的方向。

图1 基于韧皮部的运输和生长素极性运输的化学渗透模型(Hélène S Robert and Jiří Friml, 2009)。(a)生长素通过韧皮部从源组织、嫩叶和花芽到根和茎尖的分布。(b)化学渗透模型,基于质外体(pH 5.5)和细胞质(pH 7.0)的pH差异。质子化生长素未解离吲哚-3-乙酸(IAAH)可通过脂质膜扩散或由AUX1/LAX内流载体运输进入细胞。在细胞质中,它游离并以去质子化的形式被困于细胞内(IAA-)。IAA-可通过PGP-或PIN-型外排载体的作用排出细胞。载体的极性细胞定位决定了生长素在细胞间流动的方向。(c)质子化IAAH的结构。
介绍到这里,关于生长素的内容小远就不再细讲了,大家可能会问不是讲“追光”植物吗?怎么不解释植物为什么向光生长,背后的机制是什么?其实介绍了这么多大家自己应该也能理解个七七八八了,前面已经介绍过植物向光生长主要是因为生长素的不对称分布导致的,而生长素为什么会不对称分布,这主要涉及到植物内部的运输系统,长距离运输与短距离运输,在短距离运输中要是与方向有关就存在转运蛋白的极性亚细胞定位,这才是小远想为大家介绍的,对于定向运输,极性定位是第一步,第二步就涉及到转运蛋白的内流与外排功能,两步同时满足,植物才能将某个物质按照一定的方向进行运输,这样看来植物是真的非常聪明啊!那么下面就和小远一起去看看这两方面的内容吧!
4、极性亚细胞定位
在“A polarly localized transporter for efficient manganese uptake in rice”一文中,作者研究了水稻的锰转运蛋白MTP9的功能,在验证它是转运蛋白以及具有转运活性这一点上做了一些实验。
为了观察MTP9的组织和细胞定位,作者使用MTP9抗体进行免疫染色。结果表明,MTP9同时定位于野生型水稻主根和侧根的外皮层和内皮层(图2a、b、e、f)。在敲除突变体中,除自身荧光外,未检测到任何信号(图2c、d、g、h)。此外,MTP9在主根和侧根的两个细胞层的近端均表现出极性定位(图2b,f)。
作者进一步使用Nramp5(另一个锰转运蛋白)和MTP9的抗体进行双免疫染色。结果显示,Nramp5定位于远端,而MTP9定位于侧根的外皮层和内皮层的近端(图2i,j)。

图2 MTP9在主根和侧根定位的组织和细胞特异性(Ueno et al., 2015)。(a-d)野生型水稻(a,b)和mtp9-1突变体(c,d)主根MTP9蛋白免疫染色(红色)。(e–h)野生型水稻侧根(e,f)和mtp9-1突变体(g,h)的MTP9蛋白免疫染色(红色)。b,d,f,h分别为a,c,e,g中黄色方框的放大图像。(i,j)野生型水稻侧根MTP9蛋白(红色)与Nramp5蛋白(绿色)共同免疫染色。蓝色表示细胞壁自发荧光。j显示了i中黄色框的放大图像。en,内皮层;ex,外皮层;ep,表皮;sc,厚壁组织;pe,中柱鞘;xv,木质部导管。
为了确定MTP9的亚细胞定位,作者在水稻叶片原生质体中瞬时表达了一个GFP-MTP9融合基因。表达融合基因的细胞在叶绿体外的细胞外周和一些小泡处检测到荧光(图3a,d)。相比之下,仅用GFP转化的细胞的胞质和细胞核中都可以观察到荧光(图3e,h)。用DAPI(4',6-二脒基-2-苯基吲哚)对细胞核和MTP9抗体进行双染色显示,MTP9定位于细胞的最外层区域,而不是细胞核(图3i)。此外,蔗糖梯度(不连续)的Western blot结果显示,在相同部分的H -ATP酶中检测到MTP9(图3j)。所有这些结果一致表明,MTP9是一种质膜定位蛋白。

图3 水稻MTP9的亚细胞定位结果(Ueno et al., 2015)。(a-h)在水稻叶片制备的原生质体中瞬时表达GFP-MTP9融合蛋白(a-d)和GFP (e-h)的定位。(a,e)GFP荧光图像;(b,f)叶绿体荧光;(c,g)明场;(d,h)三个通道的组合图像。(i)侧根MTP9与细胞核双染色。红色代表抗体信号,青色代表DAPI和细胞壁自身荧光信号。箭头指示DAPI染色细胞核。(j)Western blot分析采用不连续蔗糖梯度制备的膜组分中的MTP9。H -ATP酶和V-ATP酶分别作为质膜和液泡膜的标记物。
小远叨叨
1、转运蛋白一般定位在膜上,这个大家都比较熟悉,研究它的亚细胞定位用到的方法与其它基因也没什么差别,转化原生质体或注射烟草的方式都可以,但是如果你想观察它的极性定位,这个时候就要用野生型植株或稳定转化的植株,在植物体里面观察它在细胞上的定位,这样才能得到我们想要的结果,在普通的亚细胞定位实验中我们是观察不到极性定位的!2、作者在这篇文献中观察MTP9的组织与细胞定位时,并没有用转基因植株,而是开发了MTP9的抗体进行免疫染色,这种做法在动物中应用的比较多,植物中一般很少有人这样研究,除非是一些大家研究的比较多的基因才会有对应的抗体,大多数时候都是目的基因融合荧光蛋白观察亚细胞定位,这个大家要清楚哦!
5、转运活性
5.1、蛋白脂质体体系
为了研究MTP9的金属转运活性,在大肠杆菌中过表达MTP9的cDNA。纯化后的蛋白部分显示出一个主要的蛋白条带,预期表观分子质量为74kDa(图4a)。用抗6×His和抗MTP9的抗体进行Western blot实验,进一步证实了该蛋白(图4a)。接着作者研究了这些转运体在*为蛋白质脂质体时是否具有锰的转运活性。当不包括MTP9时,没有发现转运活性。然而,含有纯化蛋白组分作为唯一蛋白质来源的蛋白质脂质体吸收了放射性锰。与含有MTP9蛋白的蛋白质脂质体进行孵化会显示通过施加人工pH梯度(酸性范围内)摄取一定量的锰,而这一pH梯度被羰基氰化物间氯苯腙(CCCP)敏感的质子导体抑制(图4b)。非放射性标记金属如锌、铁、镁和钙的存在不影响锰的吸收,而锰的存在降低了放射性标记锰的吸收(图4c)。这些结果表明,质膜上的MTP9对Mn2 具有外排转运活性。

图4 含纯化MTP9的蛋白质脂质体运输锰的特性研究(Ueno et al., 2015)。(a)MTP9的纯化。纯化后的组分(10μg)用10% SDS-PAGE分析,CBB染色显示(左)。分别用抗6×His抗体(中)和抗MTP9抗体(右)对纯化后的MTP9组分(10μg)和阴性对照膜组分(Control, 10μg)进行Western blot分析。标记蛋白的位置显示在左侧。*蛋白的位置用箭头表示。(b)含纯化MTP9的蛋白质脂质体的H /锰共运输。蛋白质脂质体在pH 5.6的缓冲液中制备,在pH5.6(白色柱子,-)、pH 7.0(黑色柱子, )或pH 7.0加2mCCCP(灰色柱子)的缓冲液中孵育1分钟后测定。(c)顺式抑制锰转运1分钟。在50M浓度下,在列出的化合物不存在或存在的情况下,测量了MTP9介导的1μM锰的转运。
注:脂质体(Liposome)是一种人工膜。在水中磷脂分子亲水头部插入水中,脂质体疏水尾部伸向空气,搅动后形成双层脂分子的球形脂质体,直径25~1000nm不等。
作者后面还用酵母突变株再次验证了MTP9对锰的转运活性,但在这里就不做详细介绍了,因为在后面的文献中会详细介绍到哦!
在“Diel magnesium fluctuations in chloroplasts contribute to photosynthesis in rice”一文中,作者对Mg转运蛋白OsMGT3的转运活性用了3种方法进行研究,方法都比较经典,想要了解就继续往下看吧!
5.2、细菌体系
为了研究OsMgT3是否具有转运Mg2 的能力,作者首先使用异丙基硫半乳糖苷(IPTG)诱导启动子将OsMGT3引入鼠伤寒沙门氏菌菌株MM281中。由于MM281缺乏三个Mg2 转运基因(CorA, MgtA和MgtB),它不能在含有少于1mM Mg的培养基中存活(Li et al., 2001)。选取拟南芥(Li et al., 2001)中已知的Mg2 转运蛋白基因AtMGT10作为阳性对照。在高Mg(1mM)存在的情况下,携带空载体EV、AtMGT10或OsMGT3的菌株增殖速度相同(图5a,b)。然而,在低Mg(0.1mM)的存在下,携带OsMGT3或AtMGT10的菌株的增殖明显优于携带EV的菌株,而携带EV的菌株的增殖没有明显的变化(图5a,b)。此外,短期动力学实验表明,表达OsMGT3的菌株在每个不同的25Mg浓度下的净摄取25Mg (Δ25Mg)远远高于含有EV的菌株(图5c)。以上结果表明,OsMGT3在细菌中具有转运Mg2 的能力。
5.3、酵母突变株体系
OsMGT3也被引入酵母突变株CM66中,该突变株由于Mg转运基因ALR1和ALR2失活从而对Mg缺乏敏感(Li et al., 2001)。与细菌的结果一致,在低Mg存在下,携带OsMGT3的CM66比携带EV的酵母生长得快得多(4mM;图5d,e)因此,25Mg剂量依赖性实验表明,携带OsMGT3的酵母对Δ25Mg的摄取远高于携带EV的酵母(图5f),表明OsMGT3在酵母中具有Mg内流活性。
5.4、非洲爪蟾卵母细胞体系
最后,将OsMGT3引入非洲爪蟾卵母细胞,进一步验证其内流活性,并使用稳定同位素25Mg研究其外排潜能。亚细胞定位结果显示,OsMGT3被错定位在卵母细胞的质膜上(图5g)(原文研究其亚细胞定位于叶绿体膜上)。与细菌和酵母的结果一致,携带OsMGT3或OsMGT3-GFP的卵母细胞比阴性对照具有更高的Δ25Mg摄取活性(图5h)。相比之下,OsMGT3在卵母细胞中没有表现出25Mg的外排活性(图5i)。综上所述,本研究的结果为OsMGT3是Mg2 的内流转运体提供了强有力的证据

图5 OsMGT3的Mg转运测定(Li et al., 2020)。(a-f)通过OsMGT3对细菌突变体MM281 (a-c)和酵母突变体CM66 (d-f)进行互补。图a和图d显示MM281 (a)和CM66 (d)的生长情况;b和e分别为MM281 (b)和CM66 (e)的增殖曲线。将含有OsMGT3、AtMGT10或EV的菌株在含有0.1或1mM MgCl2的N最小培养基上37℃培养36h,或在液体中培养,在600nm(OD600)动态监测吸收值。含OsMGT3或EV的酵母菌株在含4或100mM MgCl2的SD-尿嘧啶培养基上30℃培养3d,或在液体中培养,动态监测OD600。从OD600=1.0开始连续稀释4次,进行点板(a,d)。图(c)和(f)显示MM281 (c)和CM66 (f)中25Mg的摄取动力学。转化后的细菌和酵母菌在不同浓度的25Mg液体中培养30min。(g)卵母细胞中OsMGT3-GFP的亚细胞定位。(h)卵母细胞中Mg内流试验。将卵母细胞暴露于含有30mM 25Mg的MBS溶液中3h,然后用HNO3 洗涤和消化。(i)卵母细胞中Mg外排试验。将卵母细胞注射含50mM 25Mg的50nl溶液,转移到500μl新鲜的无Mg的 MBS(pH 6.0)中2h。2h后收集外溶液,ICP-MS测定25Mg。
小远叨叨
这里介绍的4种证明转运蛋白转运活性的方法都是很经典的方法,如果将来的某一天你研究的也正好是与此相关的内容,可以借鉴这几种方法哦!另外,小远想要说明一下,这些方法都只能在一定程度上证明它们的转运活性以及内流外排功能,植物体内这些转运蛋白是否发挥相应的功能还不能完全确定,想要得到一个确切的结果,就需要进一步验证,这个时候需要用到植物体本身,例如测定植物体不同部位某个物质的浓度,根据结果推动转运蛋白是否有相应的功能,科研需要严谨,大家最好不要根据一个实验方法得到的结果就下结论哦!
小结
小远由植物的向光性引出了转运蛋白的极性定位以及转运活性研究,之所以用生长素为大家介绍那么多背景知识,是因为非常经典,后面用营养元素转运蛋白为大家介绍具体的方法也是因为方法非常经典,并且实验图片还非常好看。小远想将这两个非常经典的东西介绍给大家,希望对大家的学习与科研有所帮助,另外,这一块的内容其实还值得深挖,但因篇幅所限小远在此就不再详细介绍了,接下来就留给大家去发挥吧!
References:
Friml J, Vieten A, Sauer M, et al. Efflux-dependent auxin gradients establish the apical–basal axis of Arabidopsis[J]. Nature, 2003, 426(6963): 147-153.
Friml J, Yang X, Michniewicz M, et al. A PINOID-dependent binary switch in apical-basal PIN polar targeting directs auxin efflux[J]. Science, 2004, 306(5697): 862-865.
Heisler M G, Ohno C, Das P, et al. Patterns of auxin transport and gene expression during primordium development revealed by live imaging of the Arabidopsis inflorescence meristem[J]. Current biology, 2005, 15(21): 1899-1911.
Li L, Tutone A F, Drummond R S M, et al. A novel family of magnesium transport genes in Arabidopsis[J]. The Plant Cell, 2001, 13(12): 2761-2775.
Li J, Yokosho K, Liu S, et al. Diel magnesium fluctuations in chloroplasts contribute to photosynthesis in rice[J]. Nature Plants, 2020, 6(7): 848-859.
Mockaitis K, Estelle M. Auxin receptors and plant development: a new signaling paradigm[J]. Annual review of cell and developmental biology, 2008, 24.
Raven J A. Transport of indoleacetic acid in plant cells in relation to pH and electrical potential gradients, and its significance for polar IAA transport[J]. New Phytologist, 1975, 74(2): 163-172.
Reinhardt D. Phyllotaxis—a new chapter in an old tale about beauty and magic numbers[J]. Current opinion in plant biology, 2005, 8(5): 487-493.
Robert H S, Friml J. Auxin and other signals on the move in plants[J]. Nature chemical biology, 2009, 5(5): 325-332.
Rubery P H, Sheldrake A R. Carrier-mediated auxin transport[J]. Planta, 1974, 118(2): 101-121.
Sabatini S, Beis D, Wolkenfelt H, et al. An auxin-dependent distal organizer of pattern and polarity in the Arabidopsis root[J]. Cell, 1999, 99(5): 463-472.
Ueno D, Sasaki A, Yamaji N, et al. A polarly localized transporter for efficient manganese uptake in rice[J]. Nature plants, 2015, 1(12): 1-8.
Vanneste S, Friml J. Auxin: a trigger for change in plant development[J]. Cell, 2009, 136(6): 1005-1016.
NO.1好文推荐Historical articles
“追光”植物背后的故事(一)
植物内部的“飞鸽传书”是如何实现的?
NO.2伯远生物可提供以下技术服务及试剂Technical service
分子生物学载体构建
十七大植物遗传转化
各物种基因编辑全套服务
分子检测业务
蛋白-蛋白、蛋白-核酸相互作用筛选及验证
伯远严选试剂商城-伯远严选,为你而选!
,