第一章 蛋白质的结构与功能
第一节 蛋白质的分子组成
一、蛋白质的主要组成元素:C、H、O、N、S 特征元素:N(16%) 特异元素:S
凯氏定氮法:每克样品含氮克数×6.25×100=100g样品中蛋白质含氮量(g%)
组成蛋白质的20种氨基酸
(名解)不对称碳原子或手性碳原子:与四个不同的原子或原子基团共价连接并因而失去对称性的四面体碳
为L-α-氨基酸,其中脯氨酸(Pro)属于L-α-亚氨基酸
不同L-α-氨基酸,其R基侧链不同
除甘氨酸(Gly)外,都为L-α-氨基酸,有立体异构体
组成蛋白质的20种氨基酸分类
非极性氨基酸:甘氨酸(Gly)、丙氨酸(Ala)、缬氨酸(Val)、
亮氨酸(Leu)、异亮氨酸(Ile)、脯氨酸(Pro)
极性中性氨基酸:丝氨酸(Ser)、半胱氨酸(Cys)、蛋氨酸(Met)
天冬酰胺(Asn)、谷氨酰胺(Gln)、苏氨酸(Thr)
芳香族氨基酸:苯丙氨酸(Phe)、色氨酸(Trp)、酪氨酸(Tyr)
酸性氨基酸:天冬氨酸(Asp)、谷氨酸(Glu)
碱性氨基酸:赖氨酸(Lys)、精氨酸(Arg)、组氨酸(His)
其中:含硫氨基酸包括:半胱氨酸、蛋氨酸
四、氨基酸的理化性质
1、两性解离及等电点
① 氨基酸分子中有游离的氨基和游离的羧基,能与酸或碱类物质结合成盐,故它是一种两性电解质。
② 氨基酸是两性电解质,其解离程度取决于所处溶液的酸碱度。
③(名解)等电点(pI点):在某一pH的溶液中,氨基酸解离成阳离子和阴离子的趋势及程度相等,成为兼性离子,呈电中性,此时溶液的pH称为该氨基酸的等电点。
pH<pI 阳离子 氨基酸带净负电荷,在电场中将向正极移动
pH=pI 氨基酸的兼性离子
pH>pI 阴离子 氨基酸带净正电荷,在电场中将向负极移动
④ 在一定pH范围内,氨基酸溶液的pH离等电点越远,氨基酸所携带的净电荷越大
2、含共轭双键的氨基酸具有紫外吸收性质
色氨酸、酪氨酸的最大吸收峰在 280 nm 附近
大多数蛋白质含有这两种氨基酸残基,所以测定蛋白质溶液280nm的光吸收值是分析溶液中蛋白质含量的快速简便的方法
3、氨基酸与茚三酮反应生成蓝紫色化合物
在pH5~7,80~100℃条件下,氨基酸与茚三酮水合物共热,可生成蓝紫色化合物,其最大吸收峰在570nm处。此吸收峰值与氨基酸的含量存在正比关系,因此可作为氨基酸定量分析方法
五、蛋白质是由许多氨基酸残基组成的多肽链
(一)氨基酸通过肽键连接而形成肽
1、(名解)肽键(peptide bond)是由一个氨基酸的α-羧基与另一个氨基酸的α-氨基脱水缩合而形成的化学键
2、肽是由氨基酸通过肽键缩合而形成的化合物
3、10个以内氨基酸连接而成多肽称为寡肽;由更多的氨基酸相连形成的肽称多肽
肽链中的氨基酸分子因为脱水缩合而基团不全,被称为氨基酸残基
多肽链(polypeptide chain)是指许多氨基酸之间以肽键连接而成的一种结构
多肽链有两端:N末端:多肽链中有游离α-氨基的一端
C末端:多肽链中有游离α-羧基的一端
6、AA顺序从氨基端(N-端)开始以羧基端(C-端)氨基酸残基为终点
7、人体内存在多种重要的生物活性的肽:
谷胱甘肽(GSH):是由谷氨酸、半胱氨酸和甘氨酸组成的三肽。
半胱氨酸的巯基是该化合物的主要功能基团。
GSH的巯基具有还原性,可作为体内重要的还原剂保护体内蛋白质或酶分子中巯基免被氧化,使蛋白质或酶处于活性状态。
第二节 蛋白质的分子结构
一、蛋白质的分子结构
一级结构
二级结构 ↘
三级结构 → 高级结构(空间构象)
四级结构 ↗
(一)、蛋白质的一级结构:蛋白质的一级结构指在蛋白质分子中,从N-端至C-端的氨基酸排列顺序,即蛋白质分子中氨基酸的排列顺序。
1、主要化学键:肽键,有些蛋白质还包含二硫键。
2、一级结构是蛋白质空间构象和特异生物学功能的基础
(二)、蛋白质的二级结构:指蛋白质分子中某一段肽链的局部空间结构,也就是该段肽链骨架原子的相对空间位置,并不涉及氨基酸残基侧链的构象。
1、主要化学键:氢键
2、(名解)肽键平面/肽单元:参与肽键的6个原子C 、C、O、N、H、C 位于同一平面,又叫酰胺平面或肽键平面,此同一平面上的6个原子构成了所谓的肽单元
3、二级结构以一级结构为基础,多为短距离效应。可分为:
α-螺旋:多肽链主链围绕中心轴呈有规律地螺旋式上升,顺时钟走向,即右手螺旋,每隔3.6个氨基酸残基上升一圈,螺距为0.54nm。α-螺旋的每个肽键的N-H和第四个肽键的羧基氧形成氢键,氢键的方向与螺旋长轴基本平形。
影响α-螺旋形成因素:R基较大,空间位阻大。
带相同电荷的R基集中,电荷相互排斥。
多肽链中若有脯氨酸不能形成a-螺旋。
β-折叠:多肽链充分伸展,各肽键平面折叠成锯齿状结构,侧链R基团交错位于锯齿状结构上下方;它们之间靠链间肽键羧基上的氧和亚氨基上的氢形成氢键维系构象稳定.
β-转角:常发生于肽链进行180度回折时的转角上,含4个氨基酸残基,第二个残基常为脯氨酸。
无规卷曲:无确定规律性的那段肽链。
主要化学键:氢键。
4、模体(motif)是具有特殊功能的超二级结构,是由二个或三个具有二级结构的肽段,在空间上相互接近,形成一个特殊的空间构象
(三)、蛋白质的三级结构:整条肽链中全部氨基酸残基的相对空间位置,即肽链中所有原子在三维空间的排布位置。
1、主要化学键(非共价键):疏水键(最主要)、盐键(离子键)、氢键、范德华力、二硫键
2、(名解)结构域:分子量较大的蛋白质常可折叠成多个结构较为紧密的区域,并各行其功能,称为结构域
(四)、蛋白质的四级结构:蛋白质分子中各亚基的空间排布及亚基接触部位的布局和相互作用,称为蛋白质的四级结构。由一条肽链形成的蛋白质没有四级结构。
1、(名解)亚基:有些蛋白质分子含有二条或多条多肽链,每一条多肽链都有完整的三级结构,称为蛋白质的亚基,亚基之间的结合力主要是氢键和离子键
2、主要化学键:疏水键、氢键、离子键
二、蛋白质的分类
根据组成成分:单纯蛋白质
结合蛋白质 = 蛋白质部分 非蛋白质部分(辅基)
根据形状:纤维状蛋白质
球状蛋白质
第三节 蛋白质结构与功能关系
一、 蛋白质一级结构是高级结构与功能的基础
1、蛋白质一级结构是空间构象和特定生物学功能的基础。一级结构相似的多肽或蛋白质,其空间构象以及功能也相似。
尿素或盐酸胍可破坏次级键
β-巯基乙醇可破坏二硫键
2、由蛋白质分子发生变异所导致的疾病,称为“分子病”,如:镰刀形红细胞贫血症
二、蛋白质空间结构是蛋白质特有性质和功能的结构基础。
肌红蛋白:只有三级结构的单链蛋白质,易与氧气结合,氧解离曲线呈直角双曲线。
血红蛋白:具有4个亚基组成的四级结构,可结合4分子氧。成人由两条α-肽链(141个氨基酸残基)和两条β-肽链(146个氨基酸残基)组成。在氧分压较低时,与氧气结合较难,氧解离曲线呈S状曲线。因为:第一个亚基与氧气结合以后,促进第二及第三个亚基与氧气的结合,当前三个亚基与氧气结合后,又大大促进第四个亚基与氧气结合,称正协同效应。结合氧后由紧张态变为松弛态。
三、若蛋白质的折叠发生错误,尽管其一级结构不变,但蛋白质的构象发生改变,仍可影响其功能,严重时可导致疾病发生,称为蛋白质构象疾病,如:人纹状体脊髓变性病(CJD)、老年痴呆症、亨廷顿舞蹈病、疯牛病等。
第四节 蛋白质的理化性质
1、蛋白质的两性电离:蛋白质两端的氨基和羧基及侧链中的某些基团,在一定的溶液pH条件下可解离成带负电荷或正电荷的基团。
2、蛋白质的沉淀:在适当条件下,蛋白质从溶液中析出的现象。包括:
a.丙酮沉淀,破坏水化层。也可用乙醇。
b.盐析,将硫酸铵、硫酸钠或氯化钠等加入蛋白质溶液,破坏在水溶液中的稳定因素电荷而沉淀。
变性的蛋白质易于沉淀,有时蛋白质发生沉淀,但并不变性
3、蛋白质变性:在某些物理和化学因素作用下,其特定的空间构象被破坏,从而导致其理化性质的改变和生物活性的丧失。主要为二硫键和非共价键的破坏,不涉及一级结构的改变。变性后,其溶解度降低,粘度增加,结晶能力消失,生物活性丧失,易被蛋白酶水解。常见的导致变性的因素有:加热、乙醇等有机溶剂、强酸、强碱、重金属离子及生物碱试剂、超声波、紫外线、震荡等。
4、蛋白质的紫外吸收:由于蛋白质分子中含有共轭双键的酪氨酸和色氨酸,因此在280nm处有特征性吸收峰,可用蛋白质定量测定。
5、蛋白质的呈色反应
a.茚三酮反应:经水解后产生的氨基酸可发生反应,蛋白质分子中游离a-氨基能与茚三酮反应生成蓝紫色化合物
b.双缩脲反应:蛋白质和多肽分子中肽键在稀碱溶液中与硫酸酮共热,呈现紫色或红色。氨基酸不出现此反应。蛋白质水解加强,氨基酸浓度升高,双缩脲呈色深度下降,可检测蛋白质水解程度。
第五节、蛋白质的分离和纯化
1、沉淀,
2、电泳:蛋白质在高于或低于其等电点的溶液中是带电的,在电场中能向电场的正极或负极移动。根据支撑物不同,有薄膜电泳、凝胶电泳等。
3、透析:利用透析袋把大分子蛋白质与小分子化合物分开的方法。
4、层析:
a.离子交换层析,利用蛋白质的两性游离性质,在某一特定PH时,各蛋白质的电荷量及性质不同,故可以通过离子交换层析得以分离。如阴离子交换层析,含负电量小的蛋白质首先被洗脱下来。
b.分子筛,又称凝胶过滤。小分子蛋白质进入孔内,滞留时间长,大分子蛋白质不能时入孔内而径直流出。
5、超速离心:既可以用来分离纯化蛋白质也可以用作测定蛋白质的分子量。不同蛋白质其密度与形态各不相同而分开。
第二章 核酸的结构与功能
核酸:是以核苷酸为基本组成单位的生物信息大分子,携带和传递遗传信息
核算的分类及分布:
脱氧核糖核酸(DNA):存在与细胞核、线粒体
携带遗传信息,通过复制传递给下一代
基本组成单位是脱氧核苷酸
核糖核苷酸(RNA):存在于细胞核、细胞质、线粒体
DNA转录的产物,参与遗传信息的复制表达
也可作为遗传物质
基本组成单位是核糖核苷酸
第一节 核酸的化学组成及一级结构
核酸的元素组成:C、H、O、N、P 标记元素P:用于测量生物样品中核酸的含量
核酸的分子组成: 核酸(DNA和RNA)
↓ 3',5'-磷酸二酯键
核苷酸
↙ ↘ 磷脂键
磷酸 核苷和脱氧核苷
↙ ↘ 糖苷键
戊糖 碱基
↙ ↘ ↙ ↘
核糖 脱氧核糖 嘌呤 嘧啶
核苷酸是构成核酸的基本组成单位
碱基
DNA:A、G、C、T
RNA:U、G、C、T
戊糖(主要区别)
DNA:脱氧核糖
RNA:核糖
核苷/脱氧核苷:由碱基和核糖或脱氧核糖生成
核苷酸:核苷或脱氧核苷C-5'上的羟基与磷酸通过脂键结合构成
DNA:AMP、GMP、CMP、TMP
RNA:AMP、GMO、CMP、UMP
核苷酸衍生物:
环化核苷酸:细胞信号转到中的第二信使
DNA是脱氧核苷酸通过3',5'-磷酸二酯键连接形成的大分子
一个脱氧核糖核苷酸3'-羟基与另一脱氧核糖核苷酸的5'-磷酸基团缩合形成3',5'-磷酸二酯键
DAN链:多个脱氧核苷酸通过磷脂二脂键构成了具有方向性的线性分子,称为多聚脱氧核苷酸,即DNA链
DAN链的方向是5'-端向3'-端
交替的磷脂基团和戊糖构成DAN的骨架
RAN也是具有3',5'-磷酸二酯键的线性大分子
核酸的一级结构是核苷酸的排列顺序
核苷酸在多肽链上的排列顺序为核酸的一级结构,核苷酸之间通过3',5'-磷酸二酯键连接。
2、核苷酸序列:方向是5'-端向3'-端
3、碱基序列
第二节 DNA的二级结构与功能
DNA的二级结构是双螺旋结构
(一)Changaff规则:A与T的摩尔数相等,G与C的摩尔数相等
不同生物种属的DNA碱基组成不同
同一个体的不同器官、不同组织的DNA具有相同的剪辑组成
(二)DNA双螺旋结构模型的要点
1、DNA的反向平行,右手螺旋的双链结构
脱氧核糖和磷酸基团组成亲水性骨架,位于双链的外侧,而疏水的碱基位于内侧
螺旋直径为2nm,螺距为3.4nm(3.54nm)
2、DNA双链之间形成了互补碱基对
碱基之间以氢键相结合,—A=T—,—G≡C—
每一个螺旋包含了10个碱基对,每两个碱基对的旋转角度为36°
每个碱基平面之间的距离为0.34nm
3、疏水作用力和氢键共同维持着DAN双螺旋结构的稳定性,尤以前者为重要
氢键、疏水性的碱基堆积力、中和核酸链上的负电荷
(三)DNA双螺旋的多样性
B-DAN:右手螺旋,92%相对湿度下
A-DAN:右手螺旋,适湿降低
Z-DAN:左手螺旋
(四)DAN的多链螺旋结构
二、DAN的高级结构(三级结构)是超螺旋结构
(一)原核生物DAN的环形超螺旋
(二)真核生物DAN高度有序和高度致密的结构
细胞周期:松散的染色质
细胞分裂期:致密的染色体
核小体:染色质的基本组成单位,由DAN和五种组蛋白共同构成
组成:DAN:约200bp
组蛋白:H1、H2A、H2B、H3、H4
八叠体:各两组H2A、H2B、H3、H4构成
(三)DAN是遗传信息的物质基础
1、基因:指DAN中的特定区段,核苷酸排列顺序决定了基因功能
2、DNA的基本功能:生物遗传信息复制的模板和基因转录的模板,它是生命遗传繁殖的物质基础,也是个体生命活动的基础。
第三节 RNA的空间结构与功能
RNA和蛋白质共同负责基因的表达和表达过程的调控
RNA分类:信使RNA(mRNA) 合成蛋白质的模板
转运RNA(tRNA) 转运氨基酸
核糖体RNA(rRNA) 核糖体的组成成分
不均一核RNA(hnRNA) 成熟mRNA的前体
小核RNA(snRNA) 参与HnRNA的剪接、转运
一、mRNA是蛋白质合成的模板(半衰期最短)
1、hnRNA为mRNA的初级产物,经过剪接切除内含子,拼接外显子,成为成熟的mRNA并移位到细胞质
2、成熟mRNA由编码区和非编码区构成
3、真核生物mRNA: 3'-末端:多聚A尾结构
5'-末端:帽结构(7-甲基鸟嘌呤及三磷酸鸟苷)加速翻译起始速度,增强mRNA的稳定性
4、mRNA的种类最多,代谢速度最快
5、mRNA功能:把核内DNA的碱基顺序,按照碱基互补的原则,抄录并转送至胞质,以决定蛋白质合成的氨基酸排列顺序
6、mRNA分子上每3个核苷酸为一组,决定肽链上某一个氨基酸,为三联体密码。
二、tRNA是蛋白质合成的氨基酸载体(分子量最小)
1、tRNA分子中含有稀有碱基
2、二级结构为三叶草形,具有茎环结构
三环四臂:DHU环、TψC环、反密码子环;
DHU臂、TψC臂、反密码子臂、氨基酸臂
3、所有tRNA:3'-末端:CCA-OH结构
5'-末端:游离磷酸
4、tRNA三级结构为倒L型。
三、以rRNA为组分的核糖体是蛋白是合成的场所(含量最多)
1、核糖体:rRNA与核糖体蛋白共同构成核糖体,是蛋白质生物合成的场所
2、原核生物的rRNA 真核生物的rRNA
小亚基为16S 小亚基为18S
大亚基为5S、23S 大亚基为5S、5.8S、28S
3、真核生物的18SrRNA的二级结构呈花状。
四、核酶:某些小RNA 分子具有催化特定RNA降解的活性,在RNA合成后的剪接修饰中具有重要作用,这种具有催化作用的小RNA称为核酶。
第四节 核酸的理化性质
一、核算分子具有强烈的紫外吸收260nm
嘌呤,嘧啶都含有共轭双键,因此对波长260nm左右的紫外光有较强吸收
纯DNA样品260nm/280nm比值为1.8
纯RNA样品260nm/280nm比值为2.0
DNA变性是双链解离为单链的过程
1、DNA变性:某些理化因素(温度、pH、离子强度等)会导致DNA双链互补碱基对之间的氢键的发生断裂,使双链DNA解离为单链,这种现象称为DNA变性,最常用的变性方法之一是加热
2、DNA变性只改变其二级结构,不改变它的核苷酸序列
分子量不变、Tm值不变、紫外吸收增强、粘度降低
3、解链温度或熔解温度(Tm):在解链过程中,紫外吸光度的变化ΔA260达到最大变化值的一半时所对应的温度称为DNA的解链温度或称融解温度
4、DNA分子的Tm值大小与其DNA长短以及碱基中的G+C含量相关
G+C比例越高,Tm值越高:离子强度越高,Tm值越高
变性的核酸可以复性或形成杂交双链
1、复性:变性DNA在适当条件下,两条互补链可重新恢复天然的双螺旋构象,这一现象称为复性
2、退火:热变性的DNA缓慢冷却(25°) 发生复性
热变性的DNA迅速冷却(4°) 不发生复性
3、杂交双链
核酸酶(注意与核酶区别)
1、核酸酶:是所有可以水解核酸的酶,在细胞内催化核酸的降解。
2、可分为DNA酶和RNA酶;外切酶和内切酶
3、其中一部分具有严格的序列依赖性,称为限制性内切酶。
第三章 酶
一、酶的组成
单纯酶:仅由氨基酸残基构成的酶。
结合酶:酶蛋白:决定反应的特异性;
辅助因子:决定反应的种类与性质;可以为金属离子或小分子有机化合物。
可分为辅酶:与酶蛋白结合疏松,可以用透析或超滤方法除去。
辅基:与酶蛋白结合紧密,不能用透析或超滤方法除去。
酶蛋白与辅助因子结合形成的复合物称为全酶,只有全酶才有催化作用。
二、酶的活性中心
酶的活性中心由酶作用的必需基团组成,这些必需基团在空间位置上接近组成特定的空间结构,能与底物特异地结合并将底物转化为产物。对结合酶来说,辅助因子参与酶活性中心的组成。但有一些必需基团并不参加活性中心的组成。
三、酶反应动力学
酶促反应的速度取决于底物浓度、酶浓度、PH、温度、激动剂和抑制剂等。
1、底物浓度
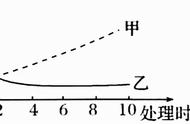
1)在底物浓度较低时,反应速度随底物浓度的增加而上升,加大底物浓度,反应速度趋缓,底物浓度进一步增高,反应速度不再随底物浓度增大而加快,达最大反应速度,此时酶的活性中心被底物饱合。
2)米氏方程式
V=Vmax〔S〕/Km+〔S〕
a.米氏常数Km值等于酶促反应速度为最大速度一半时的底物浓度。
b.Km值愈小,酶与底物的亲和力愈大。
c.Km值是酶的特征性常数之一,只与酶的结构、酶所催化的底物和反应环境如温度、PH、离子强度有关,与酶的浓度无关。
d.Vmax是酶完全被底物饱和时的反应速度,与酶浓度呈正比。
2、酶浓度
在酶促反应系统中,当底物浓度大大超过酶浓度,使酶被底物饱和时,反应速度与酶的浓度成正比关系。
3、温度
温度对酶促反应速度具有双重影响。升高温度一方面可加快酶促反应速度,同时也增加酶的变性。酶促反应最快时的环境温度称为酶促反应的最适温度。酶的活性虽然随温度的下降而降低,但低温一般不使酶破坏。
酶的最适温度不是酶的特征性常数,它与反应进行的时间有关。
4、PH
酶活性受其反应环境的PH影响,且不同的酶对PH有不同要求,酶活性最大的某一PH值为酶的最适PH值,如胃蛋白酶的最适PH约为1.8,肝精氨酸酶最适PH为9.8,但多数酶的最适PH接近中性。
最适PH不是酶的特征性常数,它受底物浓度、缓冲液的种类与浓度、以及酶的纯度等因素影响。
5、激活剂
使酶由无活性或使酶活性增加的物质称为酶的激活剂,大多为金属离子,也有许多有机化合物激活剂。分为必需激活剂和非必需激活剂。
6、抑制剂
凡能使酶的催化活性下降而不引起酶蛋白变性的物质统称为酶的抑制剂。大多与酶的活性中心内、外必需基团相结合,从而抑制酶的催化活性。可分为:
1)不可逆性抑制剂:以共价键与酶活性中心上的必需基团相结合,使酶失活。此种抑制剂不能用透析、超滤等方法去除。又可分为:
a.专一性抑制剂:如农药敌百虫、敌敌畏等有机磷化合物能特民地与胆碱酯酶活性中心丝氨酸残基的羟基结合,使酶失活,解磷定可解除有机磷化合物对羟基酶的抑制作用。
b.非专一性抑制剂:如低浓度的重金属离子如汞离子、银离子可与酶分子的巯基结合,使酶失活,二巯基丙醇可解毒。化学毒气路易士气是一种含砷的化合物,能抑制体内的巯基酶而使人畜中毒。
2)可逆性抑制剂:通常以非共价键与酶和(或)酶-底物复合物可逆性结合,使酶活性降低或消失。采用透析或超滤的方法可将抑制剂除去,使酶恢复活性。可分为:
a.竞争性抑制剂:与底物竞争酶的活性中心,从而阻碍酶与底物结合形成中间产物。如丙二酸对琥珀酸脱氢酶的抑制作用;磺胺类药物由于化学结构与对氨基苯甲酸相似,是二氢叶酸合成酶的竞争抑制剂,抑制二氢叶酸的合成;许多抗代谢的抗癌药物,如氨甲蝶呤(MTX)、5-氟尿嘧啶(5-FU )、6-巯基嘌呤(6-MP)等,几乎都是酶的竞争性抑制剂,分别抑制四氢叶酸、脱氧胸苷酸及嘌呤核苷酸的合成。
Vmax不变,Km值增大
b.非竞争性抑制剂:与酶活性中心外的必需基团结合,不影响酶与底物的结合,酶和底物的结合也不影响与抑制剂的结合。
Vmax降低,Km值不变
c.反竞争性抑制剂:仅与酶和底物形成的中间产物结合,使中间产物的量下降。
Vmax、 Km均降低
四、酶活性的调节
1、酶原的激活
有些酶在细胞内合成或初分泌时只是酶的无活性前体,必须在一定条件下,这些酶的前体水解一个或几个特定的肽键,致使构象发生改变,表现出酶的活性。酶原的激活实际上是酶的活性中心形成或暴露的过程。生理意义是避免细胞产生的蛋白酶对细胞进行自身消化,并使酶在特定的部位环境中发挥作用,保证体内代谢正常进行。
2、变构酶
体内一些代谢物可以与某些酶分子活性中心外的某一部位可逆地结合,使酶发生变构并改变其催化活性,有变构激活与变构抑制。
3、酶的共价修饰调节
酶蛋白肽链上的一些基团可与某种化学基团发生可逆的共价结合,从而改变酶的活性,这一过程称为酶的共价修饰。在共价修饰过程中,酶发生无活性与有活性两种形式的互变。酶的共价修饰包括磷酸化与脱磷酸化、乙酰化与脱乙酰化、甲基化与脱甲基化、腺苷化与脱腺苷化等,其中以磷酸化修饰最为常见。
五、同工酶
同工酶是指催化相同的化学反应,而酶蛋白的分子结构、理化性质乃至免疫学性质不同的一组酶。同工酶是由不同基因或等位基因编码的多肽链,或由同一基因转录生成的不同mRNA翻译的不同多肽链组成的蛋白质。翻译后经修饰生成的多分子形式不在同工酶之列。同工酶存在于同一种属或同一个体的不同组织或同一细胞的不同亚细胞结构中。
如乳酸脱氢酶是四聚体酶。亚基有两型:骨骼肌型(M型)和心肌型(H型)。两型亚基以不同比例组成五种同工酶,如LDH1(HHHH)、LDH2(HHHM)等。它们具有不同的电泳速度,对同一底物表现不同的Km值。单个亚基无酶的催化活性。心肌、肾以LDH1为主,肝、骨骼肌以LDH5为主。
肌酸激酶是二聚体,亚基有M型(肌型)和B型(脑型)两种。脑中含CK1(BB型);骨骼肌中含CK3(MM型);CK2(MB型)仅见于心肌。
第四章 糖代谢
一、糖酵解
1、糖酵解:在机体缺氧条件下,葡萄糖经一系列酶促反应生成丙酮酸进而还原生成乳酸的过程
2、糖酵解过程中包含两个底物水平磷酸化:一为1,3-二磷酸甘油酸转变为3-磷酸甘油酸;二为磷酸烯醇式丙酮酸转变为丙酮酸。
2、糖酵解限速酶
1)6-磷酸果糖激酶-1
变构抑制剂:ATP、柠檬酸
变构激活剂:AMP、ADP、1,6-双磷酸果糖(产物反馈激,比较少见)和2,6-双磷酸果糖(最强的激活剂)。
2)丙酮酸激酶
变构抑制剂:ATP 、肝内的丙氨酸
变构激活剂:1,6-双磷酸果糖
3)葡萄糖激酶
变构抑制剂:长链脂酰辅酶A
注:此项无需死记硬背,理解基础上记忆是很容易的,如知道糖酵解是产生能量的,那么有ATP等能量形式存在,则可抑制该反应,以利节能,上述的柠檬酸经三羧酸循环也是可以产生能量的,因此也起抑制作用;产物一般来说是反馈抑制的;但也有特殊,如上述的1,6-双磷酸果糖。特殊的需要记忆,只属少数。以下类同。关于共价修饰的调节,只需记住几个特殊的即可,下面章节提及。
一、糖酵解过程
葡萄糖
↓己糖激酶
6-磷酸葡萄糖
↓
6-磷酸果糖
↓6-磷酸果糖激酶-1
1,6-二磷酸果糖
↓
磷酸二羟丙酮
↓
3-磷酸甘油醛
↓
1,3-二磷酸甘油酸
↓
3-磷酸甘油酸
↓
2-磷酸甘油酸
↓
磷酸烯醇式丙酮酸
↓丙酮酸激酶
丙酮酸
↓
乳酸
糖酵解小结
反应部位:胞质
糖酵解是一个不需氧的产能过程。
反应全过程中有三步不可逆反应
产能方式和数量:方式:底物水平磷酸化,净产生2个ATP
二、糖有氧氧化
糖的有氧氧化(aerobic oxidation)指在机体氧供充足时,葡萄糖彻底氧化成H2O和CO2,并释放出能量的过程。是机体主要供能方式
部位:胞液及线粒体
1、过程
1)、糖酵解过程生成丙酮酸
2)、丙酮酸→→乙酰辅酶A
限速酶:丙酮酸脱氢酶复合体
辅酶有:TPP、FAD、NAD 、CoA及硫辛酸
、三羧酸循环 反应部位:线粒体
草酰乙酸+乙酰辅酶A
↓柠檬酸合成酶
柠檬酸
↓
异柠檬酸
↓异柠檬酸脱氢酶
α-酮戊二酸
↓α-酮戊二酸脱氢酶复合体
琥珀酰CoA
↓
琥珀酸
↓
延胡索酸
↓
苹果酸
↓
草酰乙酸
三羧酸循环限速酶:异柠檬酸脱氢酶
三羧酸循环关键酶:柠檬酸合酶、异柠檬酸脱氢酶、α-酮戊二酸脱氢酶
3、三羧酸循环生理意义
1)基本生理功能是氧化供能。
2)三羧酸循环是体内糖、脂肪和蛋白质三大营养物质代谢的最终共同途径。
3)三羧酸循环也是三大代谢联系的枢纽。
4、葡萄糖有氧氧化生成的32个ATP
反 应 辅酶 ATP
第一阶段 葡萄糖 6-磷酸葡萄糖 -1
三、磷酸戊糖途径生理意义
1)为核酸的生物合成提供5-磷酸核糖,肌组织内缺乏6-磷酸葡萄糖脱氢酶,磷酸核糖可经酵解途径的中间产物3- 磷酸甘油醛和6-磷酸果糖经基团转移反应生成。
2)提供NADPH
四、糖原合成与分解
1、合成过程:
葡萄糖→6-磷酸葡萄糖→1-磷酸葡萄糖→+PPi
UDPG+糖原引物→糖原
E:糖原合成酶 生成3个ATP
注:1)UDPG可看作是活性葡萄糖,在体内充作葡萄糖供体。
2、分解:肝脏。肌肉的胞浆中 关健酶:磷酸化酶
五、糖异生途径
1、 过程
乳酸 丙氨酸等生糖氨基酸
NADH
丙酮酸 丙酮酸
ATP 丙酮酸 丙酮酸
丙酮酸羧化酶
草酰乙酸 草酰乙酸 (线粒体内)
天冬氨酸 苹果酸
GTP 天冬氨酸
NADH
草酰乙酸 苹果酸
磷酸烯醇式丙酮酸羧激酶
磷酸烯醇式丙酮酸
2-磷酸甘油酸 (胞液)
ATP 3-磷酸甘油酸
NADH 1,3-二磷酸甘油酸 甘油 ATP
3-磷酸甘油醛 磷酸二羟丙酮 3-磷酸甘油
NADH
1,6-双磷酸果糖
果糖双磷酸酶
6-磷酸果糖
6-磷酸葡萄糖 1-磷酸葡萄糖 糖原
葡萄糖-6-磷酸酶
葡萄糖
注意:1)糖异生过程中丙酮酸不能直接转变为磷酸烯醇式丙酮酸,需经过草酰乙酸的中间步骤,由于草酰乙酸羧化酶仅存在于线粒体内,故胞液中的丙酮酸必须进入线粒体,才能羧化生成草酰乙酸。但是,草酰乙酸不能直接透过线粒体膜,需借助两种方式将其转运入胞液:一是经苹果酸途径,多数为以丙酮酸或生糖氨基酸为原料异生成糖时;另一种是经天冬氨酸途径,多数为乳酸为原料异生成糖时。
2)在糖异生过程中,1,3-二磷酸甘油酸还原成3-磷酸甘油醛时,需NADH,当以乳酸为原料异生成糖时,其脱氢生成丙酮酸时已在胞液中产生了NADH以供利用;而以生糖氨基酸为原料进行糖异生时,NADH则必须由线粒体内提供,可来自脂酸β-氧化或三羧酸循环。
3)甘油异生成糖耗一个ATP,同时也生成一个NADH
2、 调节
2,6-双磷酸果糖的水平是肝内调节糖的分解或糖异生反应方向的主要信号,糖酵解加强,则糖异生减弱;反之亦然。
3、 生理意义
1)空腹或饥饿时依赖氨基酸、甘油等异生成糖,以维持血糖水平恒定。
2)补充肝糖原,摄入的相当一部分葡萄糖先分解成丙酮酸、乳酸等三碳化合物,后者再异生成糖原。合成糖原的这条途径称三碳途径。
3)调节酸碱平衡,长期饥饿进,肾糖异生增强,有利于维持酸碱平衡。
第五章 脂类代谢
一、脂肪动员(fat mobilization)是指储存在脂肪细胞中的脂肪,被肪脂酶逐步水解为FFA及甘油,并释放入血以供其他组织氧化利用的过程。
脂肪动员关键酶
在脂肪动员中,脂肪细胞内的甘油三酯脂肪酶是限速酶,它受多种激素的调控,因此称为激素敏感性甘油三酯脂肪酶
脂解激素
能促进脂肪动员的激素,如肾上腺素、胰高血糖素、促肾上腺皮质激素等。
抗脂解激素
抑制脂肪动员,如胰岛素、前列腺素E2
甘油经糖代谢途径
脂酸经β-氧化分解供养
1)脂肪酸活化(胞液中)
脂酸
↓脂酰CoA合成酶
脂酰CoA(含高能硫酯键)
2)脂酰CoA进入线粒体 限速酶:肉毒碱脂酰转移酶Ⅰ
3)脂酰CoA进入线粒体基质后,进行脱氢、加水、再脱氢及硫解等四步连续反应,生成1分子比原来少2个碳原子的脂酰CoA、1分子乙酰CoA、1分子FADH2和1分子NADH。以上生成的比原来少2个碳原子的脂酰CoA,可再进行脱氢、加水、再脱氢及硫解反应。如此反复进行,以至彻底。
4)能量生成
以软脂酸为例,共进行7次β-氧化,生成7分子FADH2、7分子NADH及8分子乙酰CoA,106个ATP
三、酮体的生成和利用
组织特点:肝内生成肝外用。
合成部位:肝细胞的线粒体中。
酮体组成:乙酰乙酸、β-羟丁酸、丙酮。肝分解氧化时特有的中间产物
合成原料:乙酰CoA←脂酸氧化
限速酶:HMG CoA合酶
2、 肝外利用原因:肝内缺少乙酰乙酸硫激酶和琥珀酸 CoA转硫酶
四、脂酸的合成代谢
1、 软脂酸的合成
合成部位:线粒体外胞液中,肝是体体合成脂酸的主要场所。
合成原料:乙酰CoA、ATP、NADPH、HCO3-、Mn 等。
合成过程:限速酶:乙酰辅酶A羧化酶 辅基:生物素
激活剂:柠檬酸、异柠檬酸
甘油三酯的合成代谢
合成部位:肝、脂肪组织、小肠,其中肝的合成能力最强。
合成原料:甘油、脂肪酸
甘油一酯途径(小肠粘膜细胞)
甘油二酯途径(肝细胞及脂肪细胞)
六、甘油磷脂的合成与代谢
1、合成:肝、肾、肠内质网
2、降解
生物体内存在能使甘油磷脂水解的多种磷脂酶类,根据其作用的键的特异性不同,分为磷脂酶A1和A2,磷脂酶B,磷脂酶C和磷脂酶D。
磷脂酶A2特异地催化磷酸甘油酯中2位上的酯键水解,生成多不饱和脂肪酸和溶血磷脂。后者在磷脂酶B作用,生成脂肪酸及甘油磷酸胆碱或甘油磷酸乙醇胺,再经甘油酸胆碱水解酶分解为甘油及磷酸胆碱。磷脂酶A1催化磷酸甘油酯1位上的酯键水解,产物是脂肪酸和溶血磷脂。
七、胆固醇代谢
1、 合成
合成部位:肝是主要场所,合成酶系存在于胞液及光面内质网中。
合成原料:乙酰CoA(经柠檬酸-丙酮酸循环由线粒体转移至胞液中)、ATP、NADPH等。
合成过程:
1) 甲羟戊酸的合成(胞液中)限速酶:HMGCoA还原酶
2) 鲨烯的合成(胞液中)
3)胆固醇的合成(滑面内质网膜上)
2、去路:转化为胆汁液、转化为类固醇激素、转化为维生素D2
3、胆固醇酯的合成
细胞内游离胆固醇在脂酰胆固醇脂酰转移酶(ACAT)的催化下,生成胆固醇酯;
血浆中游离胆固醇在卵磷脂胆固醇脂酰转移酶(LCAT)的催化下,生成胆固醇酯和溶血卵磷酯。
八、血浆脂蛋白
血脂组成:甘油三酯、磷脂、胆固醇及其酯、游离脂酸
1、血浆脂蛋白分类
1)电泳法:乳糜微粒、前β、β及α
2)超速离心法:乳糜微粒(含脂最多),极低密度脂蛋白(VLDL)、低密度脂蛋白(LDL)和高密度脂蛋白(HDL),分别相当于电泳分离的CM、前β-脂蛋白、β-脂蛋白及α-脂蛋白等四类。
2、血浆脂蛋白主要由蛋白质、甘油三酯、磷脂、胆固醇及其酯组成。
乳糜微粒含甘油三酯最多,蛋白质最少,故密度最小;VLDL含甘油三酯亦多,但其蛋白质含量高于CM;LDL含胆固醇及胆固醇酯最多;含蛋白质最多,故密度最高。
血浆脂蛋白中的蛋白质部分,基本功能是运载脂类,称载脂蛋白。HDL的载脂蛋白主要为apoA,LDL的载脂蛋白主要为apoB100,VLDL的载脂蛋白主要为apoB、apoC,CM的载脂蛋白主要为apoC。
3、生理功用及代谢
1)CM 运输外源性甘油三酯及胆固醇的主要形式。成熟的CM含有apoCⅡ,可激活脂蛋白脂肪酶(LPL),LPL可使CM中的甘油三酯及磷脂逐步水解,产生甘油、脂酸及溶血磷脂等,同时其表面的载脂蛋白连同表面的磷脂及胆固醇离开CM,逐步变小,最后转变成为CM残粒。
2)VLDL 运输内源性甘油三酯的主要形式。VLDL的甘油三酯在LPL作用下,逐步水解,同时其表面的apoC、磷脂及胆固醇向HDL转移,而HDL的胆固醇酯又转移到VLDL。最后只剩下胆固醇酯,转变为LDL。
3)LDL 转运肝合成的内源性胆固醇的主要形式。肝是降解LDL的主要器官。apoB100水解为氨基酸,其中的胆固醇酯被胆固醇酯酶水解为游离胆固醇及脂酸。游离胆固醇在调节细胞胆固醇代谢上具有重要作用:①抑制内质网HMGCoA还原酶;②在转录水平上阴抑细胞LDL受体蛋白质的合成,减少对LDL的摄取;③激活ACAT的活性,使游离胆固醇酯化成胆固醇酯在胞液中储存。
4)HDL 逆向转运胆固醇。HDL表面的apoⅠ是LCAT的激活剂,LCAT可催化HDL生成溶血卵磷脂及胆固醇酯。
九、高脂血症
高脂蛋白血症分型
分型 脂蛋白变化 血脂变化
Ⅰ CM↑ 甘油三酯↑↑↑
Ⅱa LDL↑ 胆固醇↑↑
Ⅱb LDL、VLDL↑ 胆固醇↑↑甘油三酯↑↑
Ⅲ IDL↑ 胆固醇↑↑甘油三酯↑↑
Ⅳ VLDL↑ 甘油三酯↑↑
Ⅴ VLDL、CM↑ 甘油三酯↑↑↑
注:IDL是中间密度脂蛋白,为VLDL向LDL的过度状态。
家族性高胆固醇血症的重要原因是LDL受体缺陷
第七章 氨基酸代谢
一、营养必需氨基酸
简记为:缬、异、亮、苏、甲硫、赖、苯、色
二、体内氨的来源和转运
1、 来源
1)氨基酸经脱氨基作用产生的氨是体内氨的主要来源;
2)由肠道吸收的氨;即肠内氨基酸在肠道细菌作用下产生的氨和肠道尿素经细菌尿素
酶水解产生的氨。
3)肾小管上皮细胞分泌的氨主要来自谷氨酰胺在谷氨酰胺酶的催化下水解生成的氨。
2、转运
1) 丙氨酸-葡萄糖循环 肌肉到肝
2)谷氨酰胺的运氨作用 脑、肌肉等组织向肝或肾运氨
三、氨基酸的脱氨基作用
1、转氨基作用:转氨酶催化某一氨基酸的α-氨基转移到另一种α-酮酸的酮基上,生成相应的氨基酸;原来的氨基酸则转变成α-酮酸。既是氨基酸的分解代谢过程,也是体内某些氨基酸合成的重要途径。
丙氨酸转氨酶ALT 肝
天冬氨酸转氨酶 AST 心
2、L-谷氨酸氧化脱氨基作用
L-谷氨酸 L-谷氨酸脱氢酶 α-酮戊二酸+NH3
NADH
3、联合脱氨基作用
氨基酸 α-酮戊二酸 NH3+NADH
转氨酶 谷氨酸脱氢酶
α-酮酸 谷氨酸 NAD
4、嘌呤核苷酸循环
上述联合脱氨基作用主要在肝、肾等组织中进行。骨骼肌和心肌中主要通过嘌呤核苷酸循环脱去氨基。
氨基酸 α-酮戊二酸 天冬氨酸 次黄嘌呤核苷酸 NH3
GTP (IMP)
腺苷酸代琥珀酸 腺嘌呤核苷酸
(AMP)
延胡索酸
α-酮酸 L-谷氨酸 草酰乙酸
苹果酸
5、氨基酸脱氨基后生成的α-酮酸可以转变成糖及脂类,在体内可以转变成糖的氨基酸称为生糖氨基酸;能转变成酮体者称为生酮氨基酸;二者兼有者称为生糖兼生酮氨基酸。只要记住生酮氨基酸包括:亮、赖;生糖兼生酮氨基酸包括异亮、苏、色、酪、苯丙;其余为生糖氨基酸。
四、氨基酸的脱羧基作用
1、L-谷氨酸 L-谷氨酸脱羧酶 γ-氨基丁酸(GABA)
GABA为抑制性神经递质。
2、L-半胱氨酸 磺酸丙氨酸 磺酸丙氨酸脱羧酶 牛磺酸
牛磺酸是结合型胆汁酸的组成成分。
3、L-组氨酸 组氨酸脱羧酶 组胺
组胺是一种强烈的血管舒张剂,并能增加毛细血管的通透性。
4、色氨酸 色氨酸羟化酶 5-羟色氨酸 5-羟色氨酸脱羧酶 5-羟色胺(5-HT)
脑内的5-羟色胺可作为神经递质,具有抑制作用;在外周组织,有收缩血管作用。
5、L-鸟氨酸 鸟氨酸脱羧酶 腐胺 精脒 精胺
脱羧基SAM 脱羧基SAM
精脒与精胺是调节细胞生长的重要物质。合称为多胺类物质。
五、一碳单位:某些氨基酸在分解代谢过程中产生的含有一个碳原子的基团
一碳单位来源于组、色、甘、丝氨酸
一碳单位有:甲基、甲烯基、甲炔基、甲酰基及亚氨甲基,CO2不属于一碳单位。
四氢叶酸是一碳单位代谢的辅酶。
主要生理功用是作为合成嘌呤及嘧啶的原料。如N10-CHO-FH4与N5,H10=CH-FH4分别提供嘌呤合成时C2与C8的来源;N5,N10-CH2-FH4提供胸苷酸合成时甲基的来源。由此可见,一碳单位将氨基酸与核酸代谢密切联系起来。
六、芳香族氨基酸(色、酪、苯丙)的代谢
1、 苯丙氨酸
苯丙氨酸羟化酶
酪氨酸 黑色素细胞的酪氨酸酶 多巴
酪氨酸羟化酶
多巴 黑色素
多巴脱羧酶
多巴胺
SAM 去甲肾上腺素 儿茶酚胺
肾上腺素
苯酮酸尿症:当苯丙氨酸羟化酶先天性缺乏时,苯丙氨酸不能转变为酪氨酸,体内苯丙氨酸蓄积,并经转氨基作用生成苯丙酮酸,再进一步转变成苯乙酸等衍生物。此时尿中出现大量苯丙酮酸等代谢产物,称为苯酮酸尿症。
白化病:人体缺乏酪氨酸酶,黑色素合成障碍,皮肤、毛发等发白,称为白化病。
2、 色氨酸
1)生成5-羟色胺
2)生成一碳单位
3)可分解产生尼克酸,这是体内合成维生素的特例。
七、含硫氨基酸(甲硫、半胱、胱)代谢
1、甲硫氨酸 S-腺苷甲硫氨酸(SAM)
ATP PPi
SAM中的甲基为活性甲基,通过转甲基作用可以生成多种含甲基的重要生理活性物质。SAM是体内最重要的甲基直接供给体。
2、甲硫氨酸循环
甲硫氨酸 SAM 甲基转移酶 S-腺苷同型半胱氨酸
RH RCH3
甲硫氨酸合成酶 同型半胱氨酸
FH4 N5-CH3-FH4
N5-CH3-FH4可看成体内甲基的间接供体,甲硫氨酸合成酶辅酶为维生素B12。
3、肌酸的合成 肌酸以甘氨酸为骨架,由精氨酸提供脒基,SAM供给甲基而合成。在肌酸激酶催化下,肌酸转变成磷酸肌酸,并储存ATP的高能磷酸键。
4、体内硫酸根主要来源于半胱氨酸,一部分以无机盐形式随尿排出,另一部分则经ATP活化成活性硫酸根,即3'-磷酸腺苷-5'-磷酸硫酸(PAPS)。
八、氨基酸衍生的重要含氮化合物
化合物 氨基酸前体
嘌呤碱 天冬氨酸、谷氨酰胺、甘氨酸
嘧啶碱 天冬氨酸
血红素、细胞色素 甘氨酸
肌酸、磷酸肌酸 甘氨酸、精氨酸、蛋氨酸
尼克酸 色氨酸
儿茶酚胺类 苯丙氨酸、酪氨酸
甲状腺素 酪氨酸
黑色素 苯丙氨酸、酪氨酸
精胺、精脒 蛋氨酸、鸟氨酸
尿素的生成:鸟氨酸循环
1、部位:肝 线粒体、胞液
2、步奏:NH3 CO2 H2O→氨基甲酰磷酸 线粒体
E:氨基甲酰磷酸合成酶Ⅰ(CSP-Ⅰ) 耗2*ATP
N-乙酰谷氨酸
氨基甲酰磷酸 鸟氨酸→瓜氨酸 线粒体
瓜氨酸 天冬氨酸→精氨酸代琥珀酸
E:精氨酸代琥珀酸合成酶 胞液 耗ATP
精氨酸代琥珀酸→精氨酸 延胡索酸 胞液
精氨酸→尿素 鸟氨酸
尿素分子中的2个氮原子,1个来自氨,另1个来自天冬氨酸
谷氨酰胺为氮源,通过CSP-Ⅱ,催化合成氨基甲酰磷酸,并进一步参与嘧啶的合成
第八章 核苷酸代谢
一、嘌呤核苷酸代谢
合成原料:天冬氨酸 N1;甲酰基(一碳单位) C2、C8
谷氨酰胺N3 、N9;甘氨酸C5、C4、N7;CO2:C6
2、合成过程
1)从头合成:
5-磷酸核糖 PRPP合成酶 磷酸核糖焦磷酸 PRPP酰胺转移酶 5-磷酸核糖胺
ATP AMP (PRPP)
ATP AMP 次黄嘌呤核苷酸
(IMP)
GTP GMP 黄嘌呤核苷酸
(XMP)
嘌呤核苷酸是在磷酸核糖分子上逐步合成的,而不是首先单独合成嘌呤碱然后再与磷酸核糖结合而成的。
2) 补救合成:
利用体内游离的嘌呤或嘌呤核苷,经过简单的反应过程,合成嘌呤核苷酸。生理意义为:一方面在于可以节省从头合成时能量和一些氨基酸的消耗;另一方面,体内某些组织器官,如脑、骨髓等由于缺乏从头合成的酶体系,只能进行补救合成。
3、 脱氧核苷酸的生成
脱氧核苷酸的生成是在二磷酸核苷水平上,由核糖核苷酸还原酶催化,核糖核苷酸C2上的羟基被氢取代生成。
4、 分解产物
AMP 次黄嘌呤 黄嘌呤氧化酶
黄嘌呤 黄嘌呤氧化酶 尿酸
GMP 鸟嘌呤
人体内嘌呤碱最终分解生成尿酸,随尿排出体外。
痛风症患者血中尿酸含量升高。临床上常用别嘌呤醇治疗痛风症,这是因为别嘌呤醇与
次黄嘌呤结构类似,可抑制黄嘌呤氧化酶,从而抑制尿酸的生成。
,