1、基因:遗传信息的基本单位。一般指位于染色体上编码一个特定功能产物(如蛋白质或RNA分子等)的一段核苷酸序列。
2、核型:是指一个物种所特有的染色体数目和每一条染色体所特有的形态特征,包括染色体长度、着丝粒的位置、臂比值、随体的有无、次缢痕的数目及位置。
3、染色体分带:用特殊的染色方法, 使染色体产生明显的色带(暗带)和未染色的明带相间的带型, 形成不同的染色体个性, 以此作为鉴别单个染色体和染色体组的一种手段。
5、染色体带型:经过显带技术处理后的染色体,显示出特征性的带纹。每一条染色体都有固定的分带模式,即称带型。
6.操纵子:指几个功能上相近或相关的结构基因排列在一起,由一个共同的启动子、操纵子或其它调控序列来调控这些基因的转录。包括这些结构基因和控制区的整个核苷酸序列就称为操纵子。
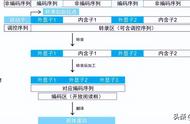
7.外显子:真核基因中与成熟mRNA、rRNA或tRNA分子相对应的DNA序列,为编码序列。
8、内含子:初级转录物中无编码意义而被切除的序列。在前体RNA中的内含子也常被称作“间插序列
9、转座子:一种复合型转座因子,这种转座因子带有同转座无关的一些基因,入抗药性基因,它的两端是插入序列,构成了“左臂”和“右臂”,两个臂可以是正向重复,也可以是反向重复。这种复合型转座因子称为转座子。
10、重叠基因:共有同一段DNA序列的两个或多个基因。重叠方式:(1)基因套基因(2)部分重叠(3)三个基因重叠
11、反转录转座子:指通过RNA为中介,反转录成DNA后进行转座的可动元件。
12、C值:一种生物单倍体基因组所含的DNA总量,称为该物种DNA的C值。
在低等真核生物中,C值的大小与生物的形态结构的复杂程度有关。而在高等生物中则不具有这一相关性。这种现象称为C值悖理。
N值悖论:处于不同进化阶梯,复杂性不同的生物种属所具有的基因数目与其结构的复杂性不成比例的现象。
13、RNP:核糖核蛋白:由RNA核糖核苷酸和蛋白质组成 。SNP: 单核苷酸的多态性:单核苷酸多态性是指在同一物种的不同个体基因组的等位序列上单个核苷酸对存在差别的现象。STS: 序列标签位点是指在染色体上定位的、序列已知的单拷贝DNA短片段。
14、转化:是指某一基因型的细胞从周围介质中吸收来自另一基因型细胞的DNA而使受体的基因型和表型发生相应变化的现象。
15、遗传*:遗传*是指一段DNA在核苷酸分子上的重新组合。
16、基因库:一个群体中所有个体的基因型的集合。
17、合胞体:昆虫的受精卵进行表面卵裂时,细胞核分裂后,细胞质并不立即发生分裂而形成的含有许多个细胞核的原生质团。
18、同源基因: 具有共同的进化起源,序列结构和功能相似的基因。
19、可变剪接:一个基因产生不止一种类型的mRNA,称为可变剪接。例如:与果蝇性别有关的基因dsx, 其mRNA在雌蝇和雄蝇中剪接方式不同。
20、基因组:单倍体细胞中整套染色体所包含的DNA分子以及DNA分子所携带的全部遗传指令。
21、假基因:来源于相应的有功能的基因,由于某种突变而失去原有的功能的基因。用Ψ表示。
22、遗传漂变: 在小群体中由于世代间配子的随机抽样造成的误差所导致的基因频率的随机波动。
23、中断杂交: 一种用来研究细菌接合生殖的实验方法。让两种菌株在培养液中混合通气培养,互相接触,形成接合管,每隔一定时间搅拌,中断接合管取样,可得到接收了不同长度的供体染色体片段的受体细菌。
24、级联反应:细胞内信号传递途径关联蛋白质的系列反应,即通过多次的逐级放大使较弱的输入信号转变为极强的输出信号,导致各种生理响应的过程。一般包括磷酸化和去磷酸化反应。
25、复等位基因:一个群体中,一对同源染色体的同一基因座上有2个以上的等位基因举例:(1)果蝇的眼色基因;(2)ABO血型系统;(3)人白细胞抗原(HLA)
26、同源*: 是指发生在姐妹染色单体之间或同一染色体上含有同源序列的DNA分子之间或分子之内的重新组合。
27、转导:通过噬菌体感染将DNA转入宿主细胞并产生新性状的过程。
普遍性转导:噬菌体能传递供体细菌的任何基因的转导。局限性转导:噬菌体只能传递供体染色体上原噬菌体整合位置附近的基因的转导。28、遗传漂变:在小群体中,由于样本的机误导致群体基因频率的随机改变,称为遗传漂变。遗传漂变的原因:奠基者效应 小群体 瓶颈效应
29、果蝇中P品系与M品系杂交中,杂种劣育现象。
(一)限制酶作图:通过比较一段DNA序列被两种限制酶切割产生的两套片段的大小,确定出酶切位点在DNA分子上的位置。
基因定位常用的方法:家系分析与基因定位(性连锁分析)、体细胞杂交【CHO与人成纤微细胞融合、(PEG、仙台病毒)、克隆分布板】、同线分析定位基因、染色体缺失定位、原位杂交、DNA序列筛查。
(二)F质粒又叫F因子,即致育因子(fertility factor)的简称,是在某些大肠杆菌细胞中发现的一种最有代表性的单拷贝的接合型质粒。F质粒有三种不同的存在方式:(i)F 细胞:以染色体外环形双链质粒DNA形式存在,其上不带有任何来自寄主染色体的基因或DNA区段。(ii)F′细胞:以染色体外环形双链质粒DNA形式存在,同时在其上还携带着细菌的染色体基因或DN区段。(iii)Hfr细胞(高频*细胞):以线性DNA形式从不同位点整合到寄主染色体。
(三)、同源*的分子机制
(一)、异源DNA双链断裂与重(错)接(二)、同源*的Holliday模型(三)、基因转变及其分子机理(四)、同源*的Meselson – Radding 模型(五) 基因转变与高度负干涉
(四)、免疫的主要过程?
1.体液免疫:A、由辅助性T淋巴细胞协助,B淋巴细胞分泌抗体;B、抗体识别并与特定的抗原结合,形成抗原-抗体复合体;C、由巨噬细胞或补体除去抗原抗体复合物。2细胞介导的免疫:A、被感染的靶细胞将抗原降解成片段;B、MHC蛋白将外来抗原片段呈递到靶细胞表面;C、Tc细胞用T细胞受体识别并去除被呈递的抗原片段。
(五)转座子类型:a. 简单转座子b.复合式转座子
转座的主要方式:a、剪-粘型转座,又叫简单插入转座,是一种非复制转座。b. 保守型转座,非复制转座的一种。c.复制型转座,在复制转座的过程中,转座子被复制,一个拷贝留在供体原位,另一个拷贝插入受体新位点。
(六)性别决定
1、 果蝇的性别决定(常染色体和性染色体套数之比决定性别、mRNA的剪接方式决定性别、sxl基因产物扳动性别决定的层次剪接)
2、 哺乳动物的性别决定:初级性决定指性腺的决定,取决于性染色体X、Y,雄性为XY,雌性为XX;次级性别决定即性别分化指性腺分泌的激素决定;Y染色体上的睾丸决定基因
3.蜜蜂中染色体倍性性别决定:2n=32发育为雌蜂;n=16,发育为雄峰。
4.后缢的位置性别决定
5爬行类的性别决定
(七)果蝇卵的极性形成、体节的分化
果蝇卵的极性形成、体节的分化是受一系列基因级联反应调控的。
参与果蝇卵的极性形成、体节的分化的基因有三类:母体效应基因(母性效应基因编码转录因子、受体和调节翻译的蛋白,在卵子发生中转录,产物储存在卵母细胞中,沿前-后轴呈梯度分布)、分节基因(分节基因决定体节的形成。主要包括:裂隙基因,成对规则基因,体节极性基因)、同源异形基因(主要包括触角足复合体和 双胸复合体)。
(八)染色体畸变
(一)染色体结构变异【1、缺失:(1) 末端缺失——猫叫综合征;(2)中间缺失:缺失的细胞遗传学效应——a. 细胞学,形成细胞环、b.遗传学;2、重复: (1)正向重复(2)反向重复;3、倒位 (1)臂内倒位(2)臂间倒位;4、易位 (1)相互易位(2)单向易位(3)罗伯逊易位。】三、染色体数目的改变 1、整倍体(草莓、西瓜加倍)2、非整倍体——21三体
(九)DNA序列的复杂性(x):最长的没有重复序列的核苷酸对数。
x=K C0t1/2
k: 与阳离子浓度、温度、DNA片段大小和 DNA序列的复杂性有关。
因此:C0t1/2也与上述因素有关。如果前三位因素确定,则C0t1/2与DNA序列的复杂性呈正相关。
(十)反转录转座:存在于真核生物中,种类复杂。根据其结构和组成,可分为两类:病毒超家族:能编码转座酶或整合酶,能自主地进行转座。如:酵母Ty转座成分、果蝇copia因子、哺乳动物,LINE1以及所有的还原病毒。非病毒超家族:自身没有转座酶或整合酶的编码能力,依靠细胞内的酶系统作用下进行转座。如:SINE等。
(十一)线虫在生物学中的贡献:
(1)特定的基因参与细胞凋亡求细胞程序性死亡的最早证据来自Rober Horritz对秀丽新小线虫发育过程的研究。
(2)线虫的细胞分裂类似于肝细胞,呈不对称分裂
(3)人体也有调控凋亡的基因,通过线虫的ced基因同源发现的线虫的ced-9基因与人体癌基因bcl-2是同源的,后者可置换秀丽新小线虫的ced-9基因。
一、自分子遗传学建立以来,人们对基因的认识有哪些修正?
1、孟德尔提出“遗传因子”概念---生物性状的遗传符号,是一些独立的遗传单位。表明生物的某些性状是由遗传因子负责传递的,遗传下来的不是形状而是遗传因子。
2、1909年,约翰逊首创了“基因”一词,用来代替孟德尔的遗传因子,并且提出基因型和表现型这样两个术语。
3、1910年,摩尔根在果蝇中发现白色复眼突变型,首先说明基因可以发生突变。
4、1926年,摩尔根提出基因的“三位一体”的概念,即基因是决定性状的最小单位、突变的最小单位、*的最小单位。
5、1928年,格里弗斯肺炎双球菌感染实验,为确定遗传物质化学性质的研究开辟了道路。
6、1953年J.D沃森和F.H.C.克里克提出了著名的DNA分子结构的双螺旋模型。
7、1957年,本泽尔提出了顺反子学说。这个学说将基因具体化为DNA分子上的一段顺序。
8、1977年,桑格等人在测定ФX174噬菌体DNA的核苷酸序列时发现:同一部分的DNA能编码两种不同蛋白质,提出了“重叠基因”的概念。两个DNA分子有一段重叠的核苷酸序列。
9、20世纪40年代,遗传学家麦克林托克在玉米研究中发现“转座因子”,说明某些基因具有游动性。50年代初麦克林托克在玉米的染色体中发现了可以改变自身位置的基因,她称之为“解离因子”。
10、操纵子进一步丰富了基因的概念,基因是可分的,不仅体现在基因的结构上,而且在功能上也可分为负责编码产生某种蛋白质分子的基因,以及负责调节其他基因功能的基因。
二、免疫球蛋白(抗体)的类型、基因组成、基因重-排及其表达方式?
1、类型:IgG、IgA、IgM、IgE、IgD。
2、基因组成:1、轻链:有κ(由Vκ、Jκ、Cκ、L基因组成)和λ(Vλ、Jλ、Cλ、L、D组成)两种——每个轻链Ig基因家族中,许多V基因片段与少得多的C基因片段相连。2、重链:每个重链Ig基因家族包括大量VH基因片段,少量D片段和J片段
3、免疫球蛋白基因重排的机制:所有片段的边界的一致序列由七聚体和九聚体组成,并由12bp或23bp的间隔序列将其分开,参与剪接反应;剪接反应的规则是具有一种间隔序列的一致序列只能与具另一类型的间隔序列的一致序列连接。
4.基因的表达:1、轻链的表达通过体细胞突变和V-C剪接;2、重链的表达通过D-J和V-D剪接。基因表达的调控:1、等位排斥:一个杂合个体的细胞只表达一对等位基因中的一个;2、类型转换:重链C基因的类型决定Ig的类型;3、增强子调节Ig基因的表达水平
三、免疫球蛋白多样性的遗传机制?
对抗体多样性的遗传控制曾提出3种学说:①种系学说(又称胚系学说):抗体形成细胞具有编码Ig分子的全部基因(即有限数量的C基因和未知数量的V基因,它是通过长期进化形成并通过生殖细胞从亲代传给子代。又称胚系学说;②体细胞突变学说:在生殖细胞内只继承了数量有限的V基因,Ig分子多样性的形成,是由于体细胞在发育过程中发生突变或基因*,从而产生许多不同的 V基因。体细胞突变可能对Ig分子的多样性发生重要作用;③V区基因相互作用学说:Ig分子可变区是由V基因片段、J基因片段和D基因片段组成,V、D、J基因相互连接对Ig分子多样性的产生,是极为重要的。Ig的多样性不可能简单地归因于上述的某一学说,它可能与上述多种机制有关,也可能还与基因片段连接点上的连接多样性以及VL和VH链的不同配对有关。
,