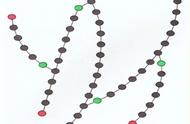
糖原(glycogen)是动物的储存多糖,主要位于肝脏和骨骼肌(肝糖原和肌糖原),脑、肾、肠中也有少量糖原。除动物外,在细菌、酵母、真菌及甜玉米中也有糖原存在。
糖原与碘反应呈紫色,光吸收在430-490nm。其结构与支链淀粉也很相似,直链部分以α-1,4糖苷键相连,分支处为α-1,6糖苷键。不过糖原分子量更大,分支密度更高。糖原分子量约为500万,端基含量占9%(支链淀粉仅4%),平均链长只有12-18个残基。

糖原结构,引自Biochem J. 2012 Feb 1;441(3):763-87.
结构必然与功能相适应。动物中的储存多糖与植物不同,淀粉主要供种子萌发时使用,而糖原代谢则随血糖和激素波动不断进行,并且需要快速响应。糖原具有更多的非还原端,所以具有更快的代谢速度。
糖原分解需要糖原磷酸化酶(glycogen phosphorylase,GP)和脱支酶(glycogen debranching enzyme,GDE)。GP在骨骼肌、肝脏和脑的同工酶分别称为GPM、GPL和GPB,编码基因分别是PYGM、PYGL和PYGB。
GP从非还原端水解α-1,4糖苷键,到分支点前4个残基停止,生成极限糊精。所以仅依靠GP只能分解糖原的40%。

糖原磷酸化反应,引自themedicalbiochemistrypage.org
GDE有两个活性中心,一个是转移酶活性(EC2.4.1.25),将外侧3个残基转移到另一条链,留下以α-1,6键相连的分支点。另一个活性中心(EC3.2.1.33)水解分支点残基,生成游离葡萄糖。

糖原脱支反应,引自themedicalbiochemistrypage.org
GP产生的葡萄糖-1-磷酸,经磷酸葡萄糖变位酶催化生成葡萄糖-6-磷酸,进入酵解、异生或磷酸戊糖途径。GDE则产生游离葡萄糖,需要耗费ATP磷酸化才能降解。所以糖原经GP水解再氧化要比游离葡糖糖酵解节省一个ATP。不过合成糖原的时候每个葡萄糖的活化反应要消耗2个高能键。
对于已有的糖原分子,其合成代谢需要糖原合酶(glycogen synthase,GS)和糖原分支酶(glycogen branching enzyme,GBE)。GS有两种同工酶,骨骼肌等大多数组织为GS1,肝脏为GS2,编码基因分别为GYS1和GYS2。
葡萄糖残基需要预先活化,反应过程与PPP相同,由UDP-葡萄糖焦磷酸化酶催化1-磷酸葡萄糖生成UDP-葡萄糖(UDPG),消耗一个UTP的两个高能键。
GS将UDPG的糖基加在糖原非还原端的C4羟基上,需要至少4个糖基的引物。所以糖原的从头合成还需要糖原蛋白(glycogenin,GN)。GN是同二聚体,由其中一个亚基将UDPG葡萄糖的C1与另一个亚基的酪氨酸羟基相连,然后陆续通过α-1,4-糖苷键形成10–20个残基的糖链,作为GS的引物。

糖原的从头合成,引自themedicalbiochemistrypage.org
与GS类似,骨骼肌等大多数组织表达GN1,肝脏则为GN2,编码基因分别为GYG1和GYG2。
分支酶由GBE1基因编码,从至少11个残基的链上将非还原端7个残基转移到较内部的位置,形成α-1,6键分支。新的分支必需与原有糖链分支距离4个残基以上。这样反复延长和分支,就形成了树状的分子。

糖原的分支反应,引自themedicalbiochemistrypage.org
糖原并非独立存在,而是与多种相关蛋白结合,形成具有不同结构和特性的糖原颗粒。糖原颗粒的结构与糖原代谢和调控密切相关,并涉及多个细胞器。下一篇文章将讨论这部分内容。